
Agelas citrina Gotera & Alcolado, 1987
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3794.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:51852298-F299-4392-9C89-A6FD14D3E1D0 |
|
DOI |
https://doi.org/10.5281/zenodo.5691145 |
|
persistent identifier |
https://treatment.plazi.org/id/03F7DF34-C01C-FFEA-FF40-CBD3DFA4EEC9 |
|
treatment provided by |
Plazi |
|
scientific name |
Agelas citrina Gotera & Alcolado, 1987 |
| status |
|
Agelas citrina Gotera & Alcolado, 1987 View in CoL
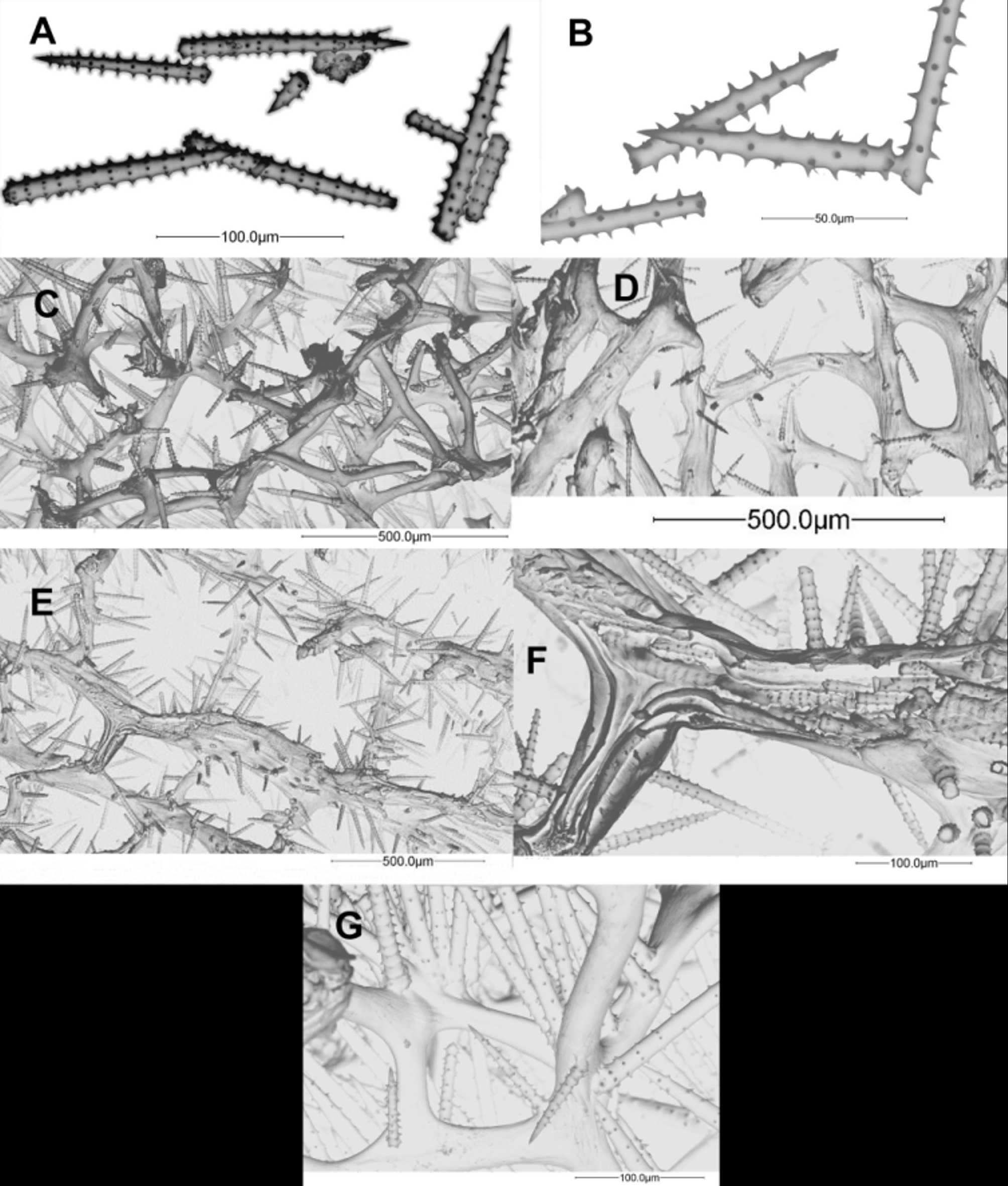
Figs. 1 View FIGURE 1 C, 1G, 13
Etymology from Latin, meaning “of citric/lemon colour”, used after the species intense yellow or orange color ( Gotera & Alcolado 1987).
Agelas citrina Gotera & Alcolado, 1987: 1 View in CoL , Figs. 1–2 View FIGURE 1 View FIGURE 2 ; Valderrama 2001: 46, Fig. 16 View FIGURE 16 ; Alcolado 2002: 61; Zea et al. 2009; Alcolado & Busutil 2012: 69.
Agelas clathrodes View in CoL ; Álvarez & Díaz 1985: 90, Fig. 26; van Soest 1981: 10; Hoppe 1988: 120, Fig. 2 View FIGURE 2 C. as A. clathrodes View in CoL 'flabelliform'; Kobluk & van Soest 1989: 1210.
Agelas View in CoL spec.; van Soest & Stentoft 1988: 99, Fig. 48.
Chalinopsis specimen labelled without specific determination, collected by Schmidt and deposited at the Zoologisk Museum København, probably collected near Caracas.
[Non: Agelas clathrodes ( Schmidt, 1870: 60) View in CoL (a valid species)]
Material and distribution. Holotype not examined, but see remarks. It is deposited at the Instituto de Oceanología — Cuba, IdO–645, collected at deep front reef southwest of Cabo Francés (west of Isla de la Juventud, Cuba), depth 40 m. The material reviewed here includes (but is not restricted to) specimens from the Bahamas (INV– POR 816, 931, 944, ICN –MHN–PO–0224), Barbados (INV– POR 916), Belize (INV– POR 951), Curaçao (INV– POR 910), Jamaica (INV– POR 791), Rosario Islands (INV– POR 969), San Andres Island (INV– POR 985, 1194); material from Los Roques (INV– POR 1198) Venezuela, was also examined (see also A. clathrodes record by Álvarez & Díaz 1985); we also examined material from Zapsurro in the southern Caribbean (INV– POR 551, described by Valderrama 2001), and San Bernardo Islands (INV– POR 1195).
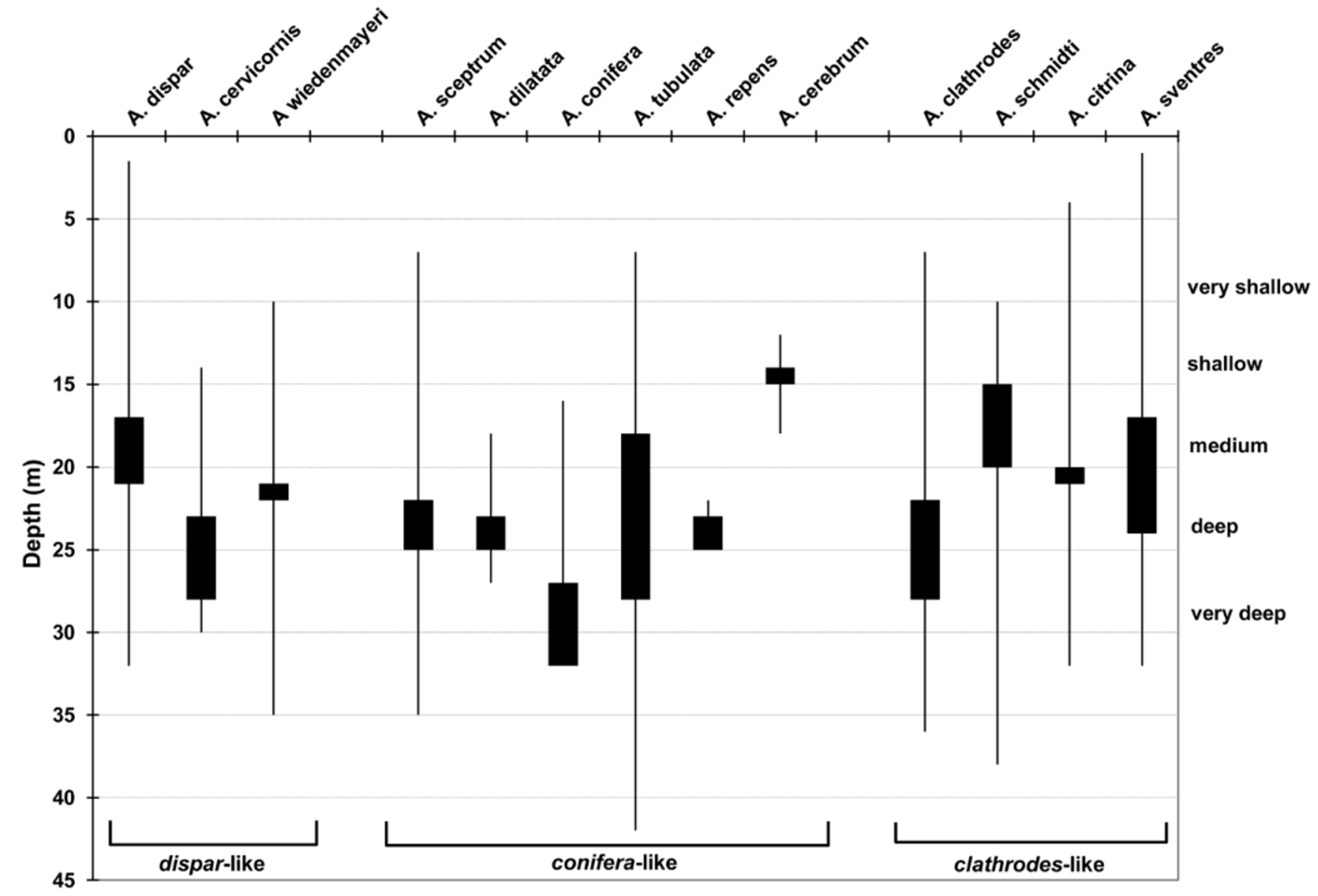
The original authors also report specimens from the same reef area at southeast of Cortés (south of Pinar del Río, Cuba), depth 60 m. S. Zea recently found it at Santa Marta, Colombia (material not included here). Besides Cuba (see also Alcolado 2002), previous records of this species are from the Bahamas and Florida Keys (Zea et al. 2009), Guadeloupe, Antigua, Barbados, Curaçao and Bonaire (van Soest 1981; Hoppe 1988; van Soest & Stentoft, 1988; Kobluk & van Soest 1989; Alcolado & Busutil 2012; Zea et al. 2009). From this information, we conclude this is a tropical northwestern Atlantic species. Our specimens were found from 4 to 32 m in depth, abundant at 20– 25 m.
Description. This species can be ear-shaped ( Fig 13 View FIGURE 13 C), lobate ( Fig 13 View FIGURE 13 H), vase-like ( Fig 13 View FIGURE 13 B), barrel-like ( Fig 13 View FIGURE 13 E) or encrusting-filling crevices ( Fig 13 View FIGURE 13 G); predominant form appears to vary geographically (see remarks). The sizes for lobate specimens are: height 5–60 cm, diameter 15–80 cm, width 2–30 cm; the sizes for barrel or irregular specimens are up to 30 cm x 40 cm. Ear-shaped specimens can reach 1 m or more in diameter in Rosario Islands. Most specimens have the external colour orange orange yellow, but also can be pink, scarlet light reddish brown, dark orange and yellow; predominant colour appears to vary geographically (see remarks), but everywhere colours have a milky appearance. Areas around the oscules or shaded areas can be milkier or white. The internal colour is orange yellow or yellow.
Clusters of 5–8 scattered oscules, often in depressions or on the inside of vases or barrels, conspicuous in most forms, but sometimes absent in fan-shape forms. Oscules are 4–12 mm wide and surrounded by a collar-like membrane; in groups of oscules the exhalant canals converge in a labyrinth fashion. Consistency in life and alcohol is strongly spongy but tough; when dry it becomes harder to cut but softer to touch. In some areas (e.g., Curaçao, Barbados, Rosario Islands, San Andres Island), specimens give off a strong smell, like rotten garlic, similar to that of Ircinia spp. The surface has regularly scattered conules, 3–6 mm height, especially in outer margins or elevated, non-oscular areas; conules are reinforced by tufts of fibres. Sometimes conules appear joined by regular ridges forming low honeycombs; sometimes the surface is irregular and contorted, with pinacoderm stretching between ridges. Pinacoderm is a bright, sometimes semitransparent, easily distinguishable membrane; it rests on nonapparent tufts of spicules protruding from main fibres. When the pinacoderm contracts, the conules are more visible and the pinacoderm can be ripped out easily. The choanosome is very cavernous; caverns lined by a bright endopinacoderm. The walls between caverns (0.3–1 cm) are dense but not firm; when macerated it is possible to see the meshes of the skeleton. The internal channels are 0.8–1.5 cm wide, and connect with the openings; in earlike specimens, the openings can perforate the entire body.
Skeletal primary fibres are 30–120 Μm in diameter, cored by 1–5 spicules per cross section, often but not always echinated; although a direction is clear, the main fibres are somewhat sinuous. Secondary fibres are sometimes echinated. Secondary and tertiary fibres are 30–40 Μm in diameter, tertiary fibres slightly narrower than secondary. The acanthostyles are straight and generally longer than in other Agelas species within several localities (see Table 2). Acanthostyles have 3–8 spines per whorl; length 88–286 (168±44.5) µm, width 4–20 (11±3.1) µm and 6–20 (13±3) whorls per spicule. Detailed lengths, widths and average number of whorls are shown in Table 2.
Remarks. Dr. P.M. Alcolado kindly checked several photographs, spicule/skeleton measurements, and descriptions of our material and agreed with our identification. This species generally grows to appreciable size on relatively calm and deep areas with medium to high coralline coverage, at the bases of coral heads or between Agaricia spp. plates; on windward, shallow-to-mid depth wave exposed settings it fills crevices in spur-and-groove systems and in the fore-reef terrace pavement.
There is regional variation in colour and shape for this species; in Venezuela and Rosario Islands the specimens are bright milky orange, massive flabellate and large, and have longer acanthostyles with more spines per whorl; in Curacao and Barbados the specimens are pink to orange, vase-like or, barrel-like; in the Bahamas most specimens are pink (a few are lemon yellow), growing as thick ears. In Curaçao and Barbados the external colour is closer to A. clathrodes but the spicule size, strong rotten garlic smell and typically conulose surface confirm they belong to A. citrina ; some difficult specimens have conules with a round aspect which could confuse the identification. Elsewhere they are predominantly chrome orange, and individuals tend to be small, crevice-filling. What Gotera & Alcolado (1987) described is thus one of the many forms, colours and consistencies that this species may take.
It was initially found to be very difficult to decide in the field whether our specimens from Curaçao, Venezuela and Barbados belonged to A. clathrodes or to A. citrina . Detailed analysis showed that they were all A. citrina (including what Álvarez & Díaz 1985 described from Los Roques, Venezuela). In reefs of Venezuela (Morrocoy, Los Roques), the often large and flabellate orange Agelas are A. citrina ; the smaller, smooth and roundish orange individuals are A. sventres ; while A. clathrodes may be rare or altogether absent (Zea, pers. obs.). At the south insular slope of Rosario Islands ( Colombia) A. citrina and A. clathrodes grow as immense ear-shaped fans that are readily distinguished by the more milky-orange appearance and rough pinacoderm of A. citrina .
A straight identification is possible wherever these two species co-exist, by spicule size and architecture. However, molecular analyses ( Parra-Velandia, 2011) showed a 10 bp deletion in the ITS1 sequence exclusive of A. citrina , which could be used to separate the most difficult specimens by means of molecular structure.
Overall, the external aspect of well-grown, erect individuals of A. citrina probably caused an earlier confusion with A. clathrodes that prevented its description as a new species until 1987, more than one hundred years after the description of A. clathrodes . Among Schmidt’s specimens at the Zoologisk Museum København kindly sent to us (including the A. clathrodes holotype), there is an additional specimen collected by Oscar Schmidt (labelled as Chalinopsis ) without a specific determination or collection site; from its spicules size, external aspect and internal architecture, that specimen, not described in any paper by Schmidt, belongs to A. citrina . So, Schmidt had the chance to compare both species and probably for reasons of prudence he did not erect a new name.
The Indonesian species A. linnaei (de Voogd et al. 2008) shares similar skeleton arrangement, spicule architecture, surface conules and chrome colour. The most remarkable difference is that choanosome of A. linnaei is dense with narrow channels. If these shared features have any evolutionary implications, they remain to be assessed.


| ICN |
Instituto de Ciencias Naturales, Museo de Historia Natural |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Agelas citrina Gotera & Alcolado, 1987
| Parra-Velandia, Fernando J., Zea, Sven & Van Soest, Rob W. M. 2014 |
Agelas
| Soest 1988: 99 |
Agelas citrina
| Alcolado 2012: 69 |
| Alcolado 2002: 61 |
| Valderrama 2001: 46 |
| Gotera 1987: 1 |
Agelas clathrodes
| Soest 1989: 1210 |
| Hoppe 1988: 120 |
| Alvarez 1985: 90 |
| Soest 1981: 10 |