
Agelas conifera ( Schmidt, 1870 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3794.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:51852298-F299-4392-9C89-A6FD14D3E1D0 |
|
DOI |
https://doi.org/10.5281/zenodo.5691131 |
|
persistent identifier |
https://treatment.plazi.org/id/03F7DF34-C013-FFD9-FF40-CBE8DBCDE8A8 |
|
treatment provided by |
Plazi |
|
scientific name |
Agelas conifera ( Schmidt, 1870 ) |
| status |
|
Agelas conifera ( Schmidt, 1870) View in CoL
Figs. 1 View FIGURE 1 A, 1E, 1F, 7, 15A
Etymology from Latin, meaning bearer of cones.
Chalinopsis conifera Schmidt, 1870: 60 View in CoL , pl. 5, Fig. 2 View FIGURE 2 b.
Synonyms and taxonomic treatment in Wiedenmayer (1977) and Zea (1987, excepting material from Santa Marta, which is here assigned to Agelas tubulata View in CoL ). In addition:
Agelas conifera View in CoL ; Álvarez & Díaz 1985: 84, Fig. 24; Kobluk & van Soest 1989: 1210; Humann 1992: 22 (in part, only right photo p. 22 and lower photo p. 23); Erhardt & Moosleitner 1995: 78 (in part, only Fig. p. 78); Gammill 1997: 15, Fig. 16 View FIGURE 16 ; Rützler et al. 2000: 235; Assmann 2000: 38, pl. 1; Assmann & Köck 2002: 157; Alcolado 2002: 61 (in part); Rützler et al. 2009: 302; Zea et al. 2009; Messing et al. 2009; Muricy et al. 2011: 38; Alcolado & Busutil 2012: 69.
Non: Agelas conifera View in CoL ; Wintermann-Kilian & Kilian 1984: 132; Zea 1987: 212 (in part, only material from Santa Marta, see pl. 13, Fig. 1 View FIGURE 1 ); Gómez 2002: 75; Díaz 2005: 470; Collin et al. 2005: 650 (= Agelas tubulata Lehnert View in CoL & van Soest, 1996).
Material and distribution. Holotype examined, deposited at the Zoologisk Museum København; it was collected in Caracas without any further information. The material reviewed here includes (but is not restricted to) specimens from the Bahamas (INV– POR 924), Belize (INV– POR 957), San Andres Island (INV– POR 977), Curaçao (INV– POR 904) and Barbados (INV– POR 912).
Other records (including accounts by Wiedenmayer 1977 and Zea 1987): Bahamas ( Gammill 1997; Assmann 2000; Zea et al. 2009), Florida East coast ( Assmann & Köck 2002; Rützler et al. 2009; Messing et al. 2009), Florida West coast in the Gulf of Mexico (Rützler et al. 2009), Cuba ( Alcolado 2002), Dominican Republic, Puerto Rico ( Weil 2006), Guadeloupe ( Alcolado & Busutil 2012), Curaçao, Bonaire (Kobluk & van Soest 1989; Zea et al.
2009), Los Roques ( Venezuela, Álvarez & Díaz 1985), Belize ( Rützler et al. 2000), and Brazil (NE in Bahia to SE in Rio de Janeiro, Muricy et al. 2011). The species has been seen by the authors in Old Providence and continental coast of Colombia, excepting Santa Marta. From the above, we consider this species as tropical western Atlantic. Its apparent absence from Santa Marta could be related to local upwelling. Our specimens were found from 11 to 42 m in depth, abundant at 25– 32 m.
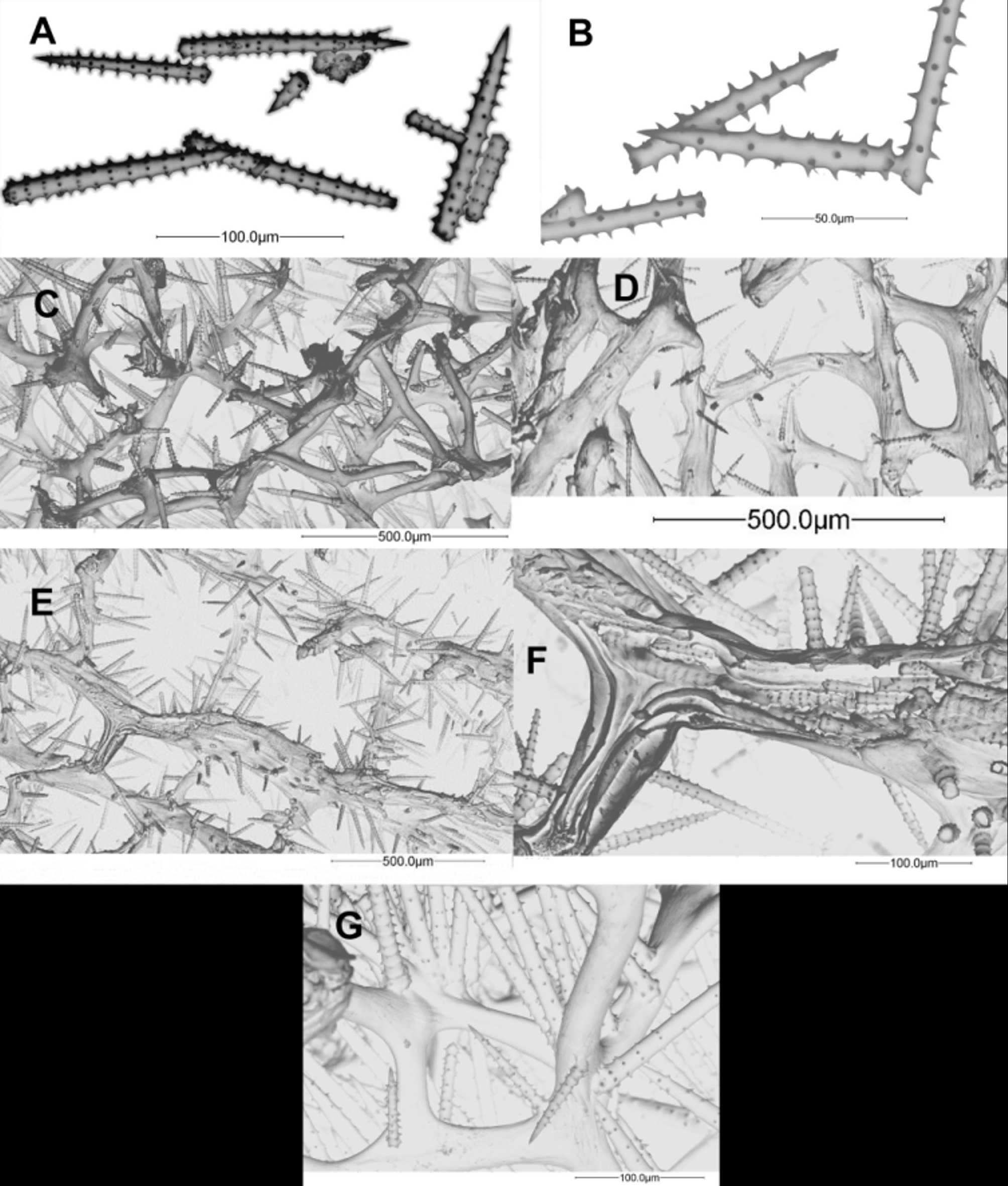
Description. This species forms an irregular flabellum or branch(es) made of oscular cones joined laterally or at their bases ( Fig 7 View FIGURE 7 A), sometimes forming repent or erect ( Fig 7 View FIGURE 7 F), elongated bodies with pinnated or alternated lobes/cones ( Fig 7 View FIGURE 7 B, 7E). When the sponge grows erect from the substratum, it may take the form of an irregular club ( Fig 7 View FIGURE 7 C). Sizes are 30–60 cm in length (but longer specimens were observed), 5–15 cm in width and 5–10 cm thick. The external colour is quite variable, bright scarlet, orange, orange yellow, brownish red, tan or even pinkish; ends and shaded parts have lighter colours, contrasting with the general specimen colour. Internal colour is quite constantly yellowish orange. Dry and preserved specimens become brownish or greyish. The pinacoderm is smooth, resting on tracts of spicules; it collapses out of the water giving a pockmarked aspect shared by A. conifera , A. tubulata , A. cerebrum and A. sceptrum .
Cones or mound openings (pseudoscula and oscula) are 0.5–3 cm in diameter, 2–4 cm deep, usually with an occluding membrane ring; tiny oscules inside atrial walls 2–5 mm in diameter. Consistency is spongy, elastic, hard to tear or cut; hard when dry. Choanosome dense, with narrow channels (<6 mm) connecting oscules.
Spongin skeleton with cored and heavily echinated primary fibres (70–110 µm in diameter) and echinated interconnecting secondary fibres (40–90 µm in diameter). Spicules are straight acanthostyles with whorls of 5–9 spines; length 72–205 (135±24.8) µm, width 3–18 (10±2.3) µm and 6–21 (13±2.7) whorls per spicule. Detailed lengths, widths and average number of whorls are shown in Table 2.
Remarks. The typical habit of this species is that of cones, wider than longer, growing on one another in an alternating or pinnate pattern; conforming to what is called the octopus form when cones are somewhat long, the club-shaped form when the sponge is erect and cones very low, or the antler-like form when groups of cones branch out. Octopus and club-shaped forms commonly coexist, being manifestations of the same growth pattern. Predominant morphology seems to be fixed for certain locations. For example, octopus-like, hanging or creeping branches predominate in Venezuela, Curaçao and Barbados, while club-shaped forms predominate in Belize and Jamaica, and antler-like morphotypes are common in the Bahamas; in the Rosario Islands both club and antler shapes are found together, the later often very long and developed, especially in deep southern-facing reef slopes where food is abundant.
Club-shaped specimens of A. conifera ( Fig. 7 View FIGURE 7 D) could be confused with the clavaeformis morphotypes of A. dispar ; however, those specimens could be separated with characters as more spines per spicule whorl in A. conifera than A. dispar and acne-like pinacoderm aspect in A. conifera vs. velvet-potato aspect in A. dispar .
A. conifera View in CoL coexists in some areas with Agelas tubulata View in CoL and A. cerebrum View in CoL , species with which it can be confused. Dr. P.M. Alcolado (pers comm., see Alcolado 2002) considers that these three species are the same (whose name should be A. conifera View in CoL ), the different growth forms being manifestations of local and micro environmental conditions and of the effect of tube size, wall thickness and presence of external oscula on the hydraulics of the pumping system. Although in a molecular study carried out in a parallel study ( Parra-Velandia 2011) there was not enough resolution to separate these three and other species, we decided to keep them separated owing to rather consistent morphological differences, especially in sites where they happened to coexist. A. tubulata View in CoL forms single tubes or groups of tubes (longer than wider), sometimes laterally fused, but usually arising from a common base; major tubes represent the whole length of the sponge. Laterally fused cones are common in A. conifera View in CoL but they do not arise from a common base, and instead arise repeatedly from one another, often conforming flabellated or antler-like shapes; a single cone rarely takes the length of the sponge. In some areas, such as Santa Marta, in the continental coast of Colombia, however, the only form present consists of clusters of long tubes arising from a rather wide base, some of them branching from one another. As major tubes span the entire length of the sponge, we decided they belong to A. tubulata View in CoL . Gómez’s (2002) A. conifera View in CoL pictured specimen from the Gulf of Mexico is clearly Agelas tubulata View in CoL as defined here. Also those pictured in Collin et al. (2005) from Bocas del Toro, Panamá. As those few specimens of tubular Agelas View in CoL observed by S. Zea in Bocas del Toro were A. tubulata View in CoL , it is probable that the record of A. conifera View in CoL by Díaz (2005) belongs to A. tubulata View in CoL .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Agelas conifera ( Schmidt, 1870 )
| Parra-Velandia, Fernando J., Zea, Sven & Van Soest, Rob W. M. 2014 |
Agelas conifera
| Alcolado 2012: 69 |
| Muricy 2011: 38 |
| Assmann 2002: 157 |
| Alcolado 2002: 61 |
| Rutzler 2000: 235 |
| Assmann 2000: 38 |
| Gammill 1997: 15 |
| Erhardt 1995: 78 |
| Humann 1992: 22 |
| Soest 1989: 1210 |
| Alvarez 1985: 84 |
Chalinopsis conifera
| Schmidt 1870: 60 |