
Zaphanta Dyar, 1910: 85
|
publication ID |
https://doi.org/10.1080/00222933.2019.1634772 |
|
DOI |
https://doi.org/10.5281/zenodo.3679845 |
|
persistent identifier |
https://treatment.plazi.org/id/03F75269-FFB4-C85B-4E9E-139CFDF9D934 |
|
treatment provided by |
Valdenar |
|
scientific name |
Zaphanta Dyar, 1910: 85 |
| status |
|
Zaphanta Dyar, 1910: 85 View in CoL
Type species. Zaphanta infantilis Dyar, 1910: 85 View in CoL .
Diagnosis
Our diagnosis follows that of Zaphantinae in St Laurent et al. (2018). Key characters, adjusted to account for the newly described species which were not examined by St Laurent et al. (2018), are as follows: Zaphanta is the only genus in which the antemedial lines on the ventral surfaces of all wings are defined. The presence of black rings of scales on the distal end of at least one pair of tibiae is unique to this genus. The remarkable external homogeneity of Zaphanta is clear in the maculation and wing shape of all Zaphanta species. Zaphanta are among the smallest Mimallonidae, with short, broad wings, yellow ground colour and purplish-pink antemedial areas on the dorsum and ventrum of all wings. ̈
Genitalia of Zaphanta are unlike any others in the family Mimallonidae ; the tegumen and uncus together form a conical structure due to the fusion of the tegumen with a usually heavily sclerotised subuncal structure. The actual shape of the subuncal structure is variable across species. A gnathos homologous with what is typically seen in other Mimallonidae genera is apparently absent. Below the uncus and subuncal sclerotisation is a variably shaped (often triangular) sclerotised plate that spans the width of the tegumen base which is present in most species. The valvae are more or less angled or curved dorsally. The phallus is variable in thickness, but usually (in all but one species) has a ventral apical extension.The female genitalia in the three species that we have examined show clear differences from each other.The ostium bursae is well sclerotised, and is distinct in appearance from the surrounding sclerotisation of segment VIII.
Description
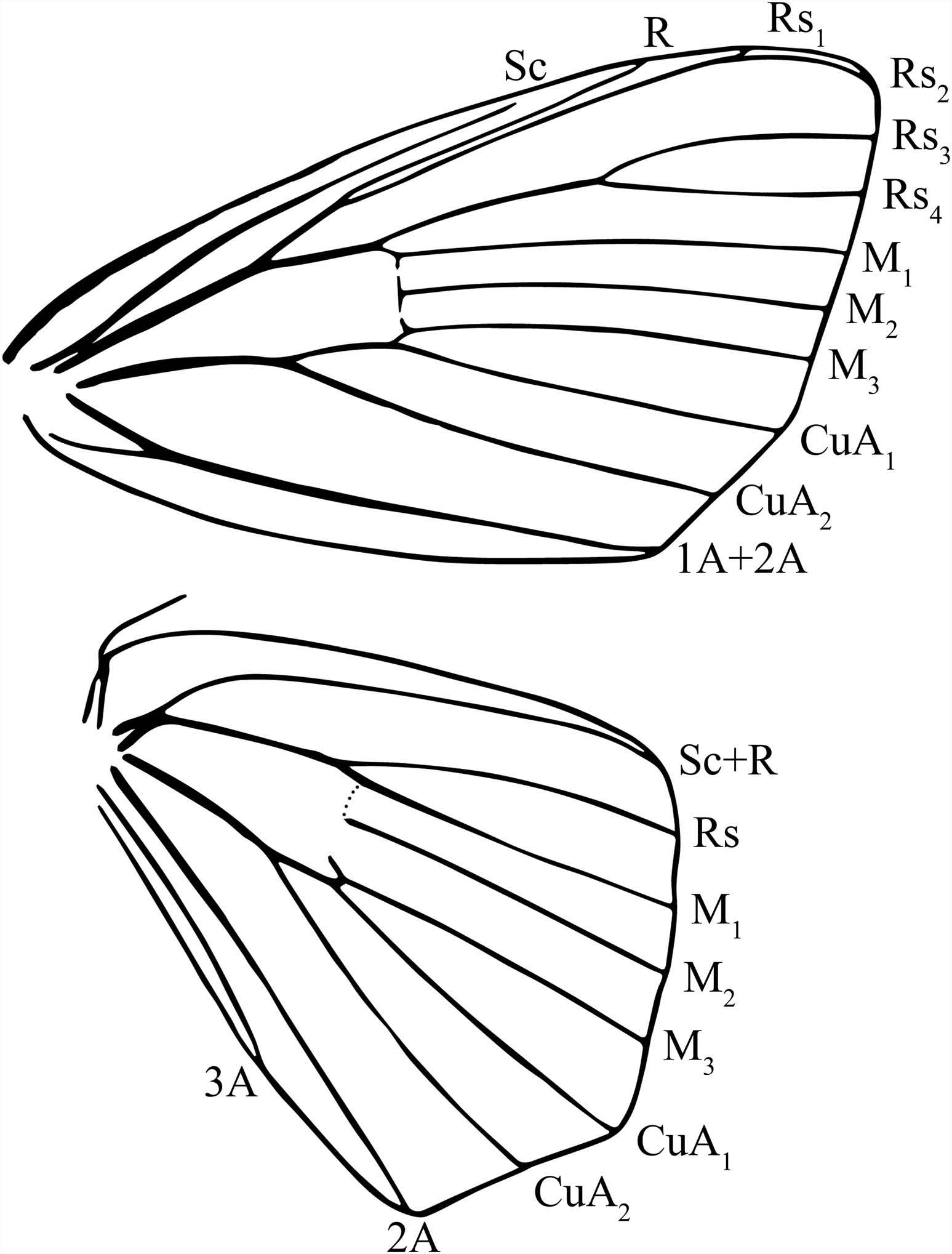
Male. Head: Light orange to yellow-brown, eyes very large, comprising more than twothirds area of head; antenna colouration as for head, antenna bipectinate to tip with pectination gradually shortening in length until tip; labial palpus reaching edge of frons, three-segmented, third segment tiny and darker scaled. Vestigial proboscis present. Thorax: Colouration as for head, rusty yellow-brown. Legs: Colouration as for thorax, except dark brown rings of scales at terminus of tibia and on terminal one to three tarsomeres. Tibial spurs, particularly mesatarsal spurs, relatively short for Mimallonidae, unapparent due to surrounding vestiture of legs. Fore wing dorsum: Fore wing length: 7.5–12.5 mm, wingspan: 17–24 mm. Somewhat rectangular in shape, apex squared or acute, margin nearly straight but angled outward approaching intersection with CuA1. Ground colour yellowish tan. Antemedial area purplish brown to pink, always distinctly darker than medial and submarginal areas. Antemedial line not apparent other than the distinct transition in colouration between ante- and medial areas. Postmedial line largely absent except for black crescent-shaped marks between veins Rs3 and M3 and occasionally between M3 and CuA2; marks more distinct nearer apex, particularly above Rs3, where marks are thicker and angle towards costa; crescent marks between M3 and CuA2 present more medially along wing, not forming continuous line with marks between Rs3 and M3. Submarginally suffused with same purplish-brown to pink colouration of antemedial area, suffusion particularly concentrated near tornus. Discal mark extremely faint to non-existent; when present appears as grey streak spanning distal expanse of discal cell. Wing overall lightly speckled by contrasting dark brown petiolate scales. Fore wing ventrum: Colouration and markings as for fore wing dorsum, but ground colour lighter overall and markings more distinct, antemedial line present as irregular black line delimiting less heavily suffused antemedial area. Discal mark and postmedial line crescents more defined than on fore wing dorsum, and present on most areas between veins. Coverage of petiolate scales generally more extensive. Hind wing dorsum: Somewhat triangular, outer margin mesally pointed. Colouration, patterning as for fore wing dorsum, but often more heavily suffused with purplish pink, particularly along entire submarginal area. Hind wing ventrum: Following same pattern as fore wing ventrum, but antemedial area weakly suffused with purplish pink. Frenulum as a single, well-defined bristle. Venation ( Figure 1 View Figure 1 ): Generally typical of Mimallonidae, but with Rs3 + Rs4 moderately to very long stalked and R + Rs1 + Rs2 all originating from the same stalk (referred to as R1–R 3 in St Laurent et al. (2018)). Abdomen: Colouration as for thorax. In well-preserved specimens, distal tip with pair of scale tufts emanating from valvae; dissection reveals that dense scale tufts originate from just above costal base of valvae. Genitalia: Relatively simple for Mimallonidae . Vinculum ovoid or somewhat rectangular. Tegumen variable, from stout to elongate. Uncus simple, triangular, often heavily sclerotised at apex, apex truncated, indented or slightly bifid. Subuncal projections quite variable, but usually heavily sclerotised, especially distally, situated immediately below uncus, usually forming a closed ring with lateral margins of tegumen; uncustegumen complex particularly elongated and narrow relative to remainder of genitalia. Mesal sclerotisation present below tegumen above valvae apodemes; apparently a component of the transtilla, this structure variable in shape. Valvae variable, but always distinctly angled or curved upwards in genital preparations (when genitalia flattened); bases of valvae meet mesally in centre of vinculum (often split apart from vinculum when genitalia flattened for photography or slide mounting); bases of valvae with accentuated saccular point that fuses to base of phallus in place of juxta. Base of valva costa variously developed; some species with valva apodeme bearing setae or spines (or an ampulla-like structure), or otherwise simple. Juxta apparently reduced to thin sclerotisation fused along basal half of phallus; juxta without ventral or dorsal components typical of Mimallonidae . Caecum of phallus present, rounded. Phallus variable in shape and length but usually with distal ventrum extended downward.
Female. Head: As for male but antennae as in male or with pectination more widely spaced and larger. Thorax: As for male. Legs: As for male. Fore wing dorsum: Fore wing length: 9–11 mm, wingspan: 22 mm. Almost identical to male, but slightly broader overall and more darkly maculated, with submarginal suffusions more distinct. Fore wing ventrum: As for male, but usually more darkly maculated with denser stippling of dark contrasting petiolate scales. Hind wing dorsum: As for male but more darkly maculated, with submarginal suffusions more distinct, contrasting against ground colour. Hind wing ventrum: As for male, but frenulum as multiple bristles. Abdomen: As for male but slightly broader. Genitalia: Tergite VIII variable in shape, region of VIII tergite proximal to base of papillae anales bilobed, weakly sclerotised, covered in short setae; sternite VIII broadened and squared. Ostium bursae globular, bowl-like, more heavily sclerotised than surrounding segment VIII. Apophyses anteriores shorter than apophyses posteriores. Ductus bursae and corpus bursae narrow, corpus bursae only slightly wider than ductus bursae, bag-like. Papillae anales irregularly shaped, distally splayed, apical angles of papillae anales extended as triangular points, basally pointed, dorsal anterior base of papillae anales with pair of small lobes densely covered in setae.
Remarks
Zaphanta are among the smallest Mimallonidae, and are poorly represented in natural history collections. We believe the rarity of Zaphanta specimens in collections may be due to the uncharacteristically geometrid-like appearance of these moths, such that they do not look like most Mimallonidae and thus may not be correctly identified as belonging to this family.
This genus is broadly distributed from Guatemala to south-eastern Brazil. The phylogenetic relationship of Zaphanta as the sister lineage to all other Mimallonidae suggests that studies focused on this genus could offer insight into the biogeographic history and ancestral host plant associations of Mimallonidae .
Identification of Zaphanta species is difficult without dissection, and even differentiating males from females is much more difficult than in all other Mimallonidae genera due to reduced sexual dimorphism. The presence of a frenulum as a single bristle in males and as multiple bristles in females is perhaps the least ambiguous method for differentiating sexes in Zaphanta without dissection or brushing off of the scales covering the genitalia. Therefore, it is advisable to dissect specimens belonging to this genus whenever possible for accurate specific identification, especially when material is collected from new regions not covered in this study. Descriptions and re-descriptions for external characters of individual Zaphanta species do not greatly differ from the generic description we give above. For descriptions we primarily focus on some key species-specific characters, which usually pertain to the genitalia. Interestingly, in the two most common species, Z. infantilis and Z. fraterna , for which we were able to examine more specimens than all other newly described species, we observed external intraspecific variation that largely encompasses the different phenotypes related to wing shape, colouration and patterning that we have seen across the entire genus. For example, the rounded wing shape seemingly unique to Z. stiletto sp. nov. was observed in some specimens of both Z. infantilis and Z. fraterna , as was the darker suffused colouration of Z. elephanta sp. nov. Other less dramatic similarities in observations are apparent as well; therefore, we primarily rely on male genitalia to recognise the various species in this odd genus. We hope that this revision will serve as a preliminary step towards understanding the genus Zaphanta , such that additional data including life history and DNA may eventually help unravel these complexities.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |