
Dendropsophus counani, Fouquet, Antoine, Orrico, Victor Goyannes Dill, Ernst, Raffael, Blanc, Michel, Martinez, Quentin, Vacher, Jean-Pierre, Rodrigues, Miguel Trefaut, Ouboter, Paul, Jairam, Rawien & Ron, Santiago, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.4052.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:176D8B81-ED65-4A0E-A49E-894F16EEF457 |
|
DOI |
https://doi.org/10.5281/zenodo.5614922 |
|
persistent identifier |
https://treatment.plazi.org/id/03F7232C-3518-FF99-968E-38A8DB7B631F |
|
treatment provided by |
Plazi |
|
scientific name |
Dendropsophus counani |
| status |
sp. nov. |
Dendropsophus counani View in CoL sp. nov.
( Figs. 3–4 View FIGURE 3. a – c View FIGURE 4 )
Hyla brevifrons View in CoL — Duellman and Crump 1974; Lescure and Marty (2000); Ernst et al. (2005); Salducci et al. (2005) Dendropsophus brevifrons View in CoL —Ernst et al. (2006); Fouquet et al. (2007); Ernst and Rödel (2008); Ouboter and Jairam (2012); Marty and Vacher (2013)
Dendropsophus cf. brevifrons View in CoL —Dewynter et al. (2008); Fouquet et al. (2011); Orrico et al. (2013)
Holotype. MNHN 2015.0101 (field number AF1345). An adult male from “Grande Montagne Tortue”, (4.292663, -52.349539, 400 m elevation), municipality of Régina, French Guiana, collected by Antoine Fouquet, Benoit Villette, and Quentin Martinez on 26 December 2013.
Paratypes. MNHN 2015.0104–7 (field number AF1346–9) two adult males and two adult females collected with the holotype. MNHN 2015.0102-3 (field numbers AF0017, AF0935) two adult males from Montagne de Kaw (4.548056, -52.151944, 250 m elevation), municipality of Roura, French Guiana, collected by Antoine Fouquet on 17 December 2005 and the 10 February 2013, respectively. MNHN 2015.0108–10 (field numbers AF1405, AF1406, AF1413) three adult males from Camp Arataie, Reserve Naturelle des Nouragues (3.99047, -52.57662, 100 m elevation), municipality of Régina, French Guiana, collected by Antoine Fouquet and Jennifer Devillechabrolle on 17 December 2013. MNHN 2015.0113 (field number AF1417) an adult male from Saut Aikoupai (4.086343, -52.462338, 100 m elevation), municipality of Régina, French Guiana, collected by Maël Dewynter on 16 December 2013. MNHN 2015.0114 (field number AM021) an adult male from Tolenga, Inini River (3.663159, -53.928308, 150 m elevation), municipality of Maripassoula, French Guiana, collected by Maël Dewynter and Kévin Pineau on 10 February 2012. MNHN 2015.0111–2 (field numbers AF 1676–1677) two adult males from Crique Limonade (3.573170, -53.198510, 100 m elevation), municipality of Saül, French Guiana, collected by Antoine Fouquet, Elodie Courtois, and Jean-Pierre Vacher on 24 February 2014. SMNS 12009–11 (field numbers MABH 1203, 1303, 1703), two adult males and one adult female from Mabura Hill Forest Reserve, Guyana (5.155278, -58.699722, 60 m elevation), collected by Raffael Ernst on 12 May 2003 and 10 June 2003, respectively. SMNS 12012–15 (field numbers MABH 0304, 0 404, 1104, 1204), three adult males and one adult female from Mabura Hill Forest Reserve, Guyana, collected by Raffael Ernst on 19 April 2004 and 29 April 2004. MNHN 2015.0115, NZCSA 2332, MNHN 2015.0116 (field numbers AF2121, AF2198, AF2199) two adult males and one female from Sipaliwini (2.09753, -56.1472, 100 m elevation), Sipaliwini District, Suriname, collected by Antoine Fouquet and Jean-Pierre Vacher on 24 April 2014. MNHN 2015.0117–8, NZCS A2333–4, MNHN 2015.0119–22 (field numbers AF2511–2, AF2516–9, AF2532–3) seven adult males and one female from Nassau mountain (4.802620, -54.615020, 400 m elevation), Sipaliwini District, Suriname, collected by Antoine Fouquet and Rawien Jairam on 20 December 2014.
Etymology. The specific epithet refers to the utopic and short-lived “independent state of Counani ” which was founded by French settlers and existed from 1886 to 1891 at the border of what is now French Guiana and the Brazilian Amapá State. It was governed by the self-proclaimed “Gros 1er”.
Diagnosis. We assigned the new species to the genus Dendropsophus on the basis of previous ( Fouquet et al. 2007, 2011) and present molecular results ( Fig. 2 View FIGURE 2 ) and the overall similarity with other species of the genus ( Figs. 3–4 View FIGURE 3. a – c View FIGURE 4 ). Dendropsophus counani sp. nov. is diagnosed by the following combination of traits: (1) a small sized member of the D. parviceps group, (SVL 19.40–21.73 mm in 11 adult males, 22.10–24.50 mm in two females) (Table 2); (2) inconspicuous sexual dimorphism, throat with dark flecks in male vs. plain cream in female, translucid glandular nuptial pads visible under a magnifying stereoscope in adult males; (3) head short, wider than body; (4) snout truncate in dorsal and lateral views, nostrils slightly protuberant; (5) large prominent eyes (ED/ HW=0.36–0.41); (6) Tibia length 51–56% SVL; (7) Femur length 37–42 % SVL; (8) palpebral membrane pigmented on its border; (9) small tympanum ( TYD /ED = 0.33–0.41); (10) well developed axillary membrane; (11) distal subarticular tubercle on finger IV strongly bifid; (12) hands webbing formula I 2 ¯—2¯ II 2 ½—2¯ III 2 +—2¯ IV, (13) feet webbing formula I 2 ½—2 II 1 +—2¯ III 1 +—2+ IV 2 +—1+ V; (14) no inner tarsal fold, supernumerary tarsal tubercles absent; (15) heel and calcar tubercles absent; (16) fringe from postaxial edge of Finger IV prolonged into a series of tubercles on forearm; (17) cloacal opening covered by a sheath on its dorsal third; (18) in life, dorsal surfaces with a few scattered tubercles, light brown in life to gray in preservative with dark transversal markings in most cases; (19) two (sometimes only one) white subocular and two white rostral (sometimes inconspicuous) markings in the upper lip, (20) neither rostral nor canthal stripes; lateral surfaces of body with whitish and dark transversal markings, (21) no dorsolateral lines; (22) dorsal surface of thighs generally black to dark gray with 0–3 cream transversal blotches situated anteriorly (sometimes inconspicuous) in either live or preserved specimens, (23) yellow blotches on thighs or tibia absent in either live or preserved specimens; (24) belly and throat white (anterior part of chin with black blotches in males), (25) ventral surface of thighs and tibia light grey and smooth, anterior surface of belly cream and granular, less granular posteriorly; (26) flanks without dorsolateral stripes, unpigmented on the axilary region, with a dark oblique marking medially followed by one or two cream markings; (27) distal parts of the ventral surfaces of hands and feet dark grey to black; (28) iris copper; (29) Advertisement call either a short trill ( 0.33 s) or click series ( 0.45 s), trills consist of 41–50 notes and click series consists of 3–6 clicks, each with 1–3 notes, these notes have a dominant frequency 3.8–4.0 kHz with a slight frequency modulation in trills; (30) in life, tadpoles light brown dorsally and marbled with black and light brown laterally, dark pigmentation concentrated on the posterior part of the tail but the tip is mostly unpigmented at the exception of a central dark marking, the tail is long with pointed tip, no row of denticles, jaw sheath finely serrated and one ventral row of blunt marginal papillae.
See comparison with other species of the parviceps group (Table 5).
TABLE 2. Morphological measurements (abbreviations and acronyms are explained in the text) of adults of Dendropsophus counani sp. nov. and other species of the D. parviceps species group. Measurements of D. brevifons are from Duellman and Crump (1974), of D. bokermanni from Goin (1960), of D. pauiniensis from Heyer (1977), and of D. frosti from Motta et al. (2012). X = and sd = standard deviation.
D. counani D. brevifrons D. bokermanni D. pauiniensis D. frosti
M (n=18) X (sd) F (n=5) X (sd) M F F ( holotype)* M ( holotype) M X (sd) F 19.40–21.73 20.53 (0.73) 22.10–24.50 23.00 (0.95) 17.0–21.4 20.1–23.2 17.8 20.2 21.1–25.2 22.1 (0.9) 25.9 9.90–11.20 10.42 (0.34) 12.00–12.65 12.10 (0.43) 8.5 9.9 11.8–12.3 12.0 (0.2) 12.9 9.20–11.10 9.90 (0.40) 11.40–12.10 11.70 (0.30) 8.2 9.5 11.5–12.0 11.8 (0.3) 12.3 5.50–6.50 5.97 (0.27) 6.50–7.10 6.79 (0.24)
7.20–8.80 8.01 (0.42) 8.70–9.70 9.21 (0.37) 11.7 7.8 9.5–9.9 9.7 (0.2) 10.3 TD 0.70–1.00 0.86 (0.07) 0.90–1.30 1.05 (0.15)
FD 0.70–1.17 0.96 (0.13) 1.10–1.30 1.18 (0.08) 1.0–1.4 1.2 (0.2) 1.0 FiL 2.50–3.20 2.74 (0.21) 2.70–3.40 3.06 (0.38)
5.36–7.00 5.85 (0.39) 5.40–6.90 6.30 (0.58)
2.52–3.30 2.87 (0.20) 3.00–3.40 3.26 (0.15)
1.4.3–2.50 2.01 (0.30) 1.85–2.30 2.21 (0.24)
5.03–7.20 6.47 (0.57) 6.38–7.60 7.10 (0.51) 5.1 7.1–8.1 7.5 (0.4) 8.6 6.30–7.50 6.77 (0.28) 7.30–7.70 7.56 (0.15) 5.1 6.8 7.3–8.8 8.0 (0.7) 8.2 2.50–3.10 2.78 (0.17) 2.80–3.00 2.88 (0.13) 3.4–3.8 3.6 (0.2) 3.2 2.60–3.00 2.74 (0.23) 2.70–3.00 3.24 (0.25)
0.90–1.10 1.00 (0.06) 1.00–1.20 1.08 (0.11) 0.9–1.4 1.2 (0.2) 1.5 It is noteworthy that the specimen of D. bokermanni designed as holotype by Goin (1960) may in fact be immature given the smallest female measured by us from Ecuador was 23.4 mm and Duellman (1978) 22.5 mm.
Description of holotype. ( Fig. 3 View FIGURE 3. a – c ) Adult male, SVL 20.9 mm; body slender; head wider than body, wider than long, HW/HL=1.1, widest behind eyes; snout truncate in dorsal and lateral views; canthus rostralis inconspicuous, curved; loreal region concave; nostrils slightly protuberant, directed laterally. Interorbital area flat; eyes large (ED/ HW=0.3) and protuberant; palpebral membrane translucent, with brown pigmentation on its border. Tympanum small ( TYD /FD=1.4), distinct but inconspicuous dorsally, directed laterally. Supratympanic fold absent. Proximal part of upper arm thickened anteriorly and slender distally, bearing an axillary membrane reaching half of upper arm ventrally; forearm angular, enlarged laterally, with tubercles on the posteroventral edges (6 on the right arm and 8 on the left) aligned with the fringe on the postaxial edge of Finger IV; fingers short, bearing round discs; relative length of fingers I <II <IV <III; subarticular tubercles large and prominently bifid on finger IV, slightly bifid on finger III, divided on finger II, round and single on finger I, most prominent on finger I; supernumerary tubercles absent; inner metacarpal tubercle flat, elongate; outer metacarpal tubercle flat large, bifid; inconspicuous white nuptial excrescences on the dorsolateral edge of finger I; webbing formula I 2 -—2- II 2 ½—2- III 2 +— 2- IV. Hindlimb long and slender; TL/SVL = 0.5; row of tubercles instead of a proper tarsal fold; calcar and heel tubercles absent; toes bearing round discs, as large as those on fingers; relative lengths of toes I <II <V <III <IV; subarticular tubercles round; supernumerary tubercles absent; inner metatarsal tubercle flat, elongate; outer metatarsal tubercle flat, round; webbing formula I 2 ½— 2 II +1+—2- III 1 +—2+ IV 2 +—1+ V. Skin on dorsum, head, dorsal surfaces of forearms and thighs, flanks and groin with homogeneously scattered tubercles; skin strongly granular on belly and posterior side of thighs, finely granular on chest and smooth on ventral surfaces of thighs, shanks, and throat. Cloacal opening directed posteriorly at midlevel of thighs, covered by cloacal sheath dorsally; cloacal tubercles absent. Tongue cordiform, barely free behind; dentigerous process of vomers evident, in two transverse series, separated and positioned obliquely to choanae, each having three vomerine teeth; choanae large, rounded; vocal slits moderately long, extending from midlateral base of tongue, almost reaching to angle of jaws; vocal sac single, median, subgular.
In alcohol, dorsal surfaces bronze grey with a few small black flecks; loreal region dark grey, one cream subocular mark finely delimited by black lines, two small cream spots on the tip of the upper lip, a small white spot at the anterior tip of eye, upper and forearm as the back but with a dark grey chevron, tibia and tarsus as back but with two dark grey chevrons, thighs white grey anteriorly and ventrally, black with a large and a few smaller cream spots dorsally, half black half grey posteriorly, venter creamy white, ventral sides of tibia, chest and throat white grey except the chin which is cream with black flecks; iris copper.
Measurements of holotype (in mm). SVL:20.9; TiL:10.4; ThL:9.8; TaL:5.7; FL:8.3; 4TD:0.9; 3FD:0.8; 1FiL:2.7; HaL:6.0; ETS:2.9; EN:2.3; HL:7.2; HW:7.5; ED:2.8; IOD:2.9; TYD:1.0 (Table 2).
Color variation of type series. Dendropsophus counani sp. nov. has a polychromatic dorsum ( Fig. 4 View FIGURE 4 ) and, as most hylids, displays a nocturnal and a diurnal coloration ( Fig. 3 View FIGURE 3. a – c ). Given the specimens were fixed during the day the variation reported here concerns the diurnal colouration. The background colouration varies from dark brown to grey. The dorsal markings varies from a conspicuous “X” extending from the head to the flanks and a “V” extending from the posterior part of the back to the inguinal region (MNHN2015.0107), to irregular transversal marks (MNHN2015.0109) to almost absent (MNHN2015.0101). The dorsal surface of the thighs is also very variable, typically including three conspicuous cream blotches (one inconspicuous pericloacal, one largest and conspicuous central and one before the knee) elongated transversally and located on the anterior part of the dorsal surface of the thigh (MNHN2015.0108); to inconspicuous (MNHN2015.0104). The loreal region comprises one or two subocular cream blotches. The gular region is generally spotless white in female but with black flecks on the anterior part in males.
Advertisement call. The following description is based on four recordings ( Table 3 View TABLE 3 ). Three of them are from uncollected individuals from Camp Aratai (Réserve Naturelle des Nouragues, French Guiana, available in Marty & Gaucher 1999), Crique Limonade (Saül, French Guiana) and Mabura Hill forest Reserve ( Guyana) and one collected (MNHN2015.0114) from Inini ( French Guiana).
The call of Dendropsophus counani sp. nov. consists of trills and clicks series ( Fig. 5 View FIGURE 5 ). The trills are short (range = 0.21– 0.35 s), comprise 33–50 notes (range = 134–153 notes/s) and are emitted between silent intervals of 1.6– 23.9 s. These trills are generally followed by click series (range = 3–6 clicks/serie; 8.4–10.7 clicks/s) consisting of generally 2–3 notes per click. However, single click series are emitted more frequently than trills (between silent intervals of X = 3.7 s; range = 2.2– 5.4 s), and last c.a. 0.45 s (range = 0.33– 0.54 s).
The spectral structure of trills and clicks are similar and consist of three main harmonics, with the fundamental frequency dominating (dominant frequency X = 3.9 kHz, range = 3.78–3.97 kHz). Two secondary harmonics are visible, having a frequency of ca. 7.6–8.1 and 10.6–11.9 kHz, respectively ( Fig. 5 View FIGURE 5 ). See comparison with other species of the parviceps group (Table 5).
Tadpole description. The following description is based on three individuals at stage 31 ( Fig. 6 View FIGURE 6 ). Type 4 tadpole (Orton 1953); exotrophic; body skin smooth; TL 12.6–14.5 mm; BL 4.2–4.6 mm, 29–36% of TL, 1.3–1.6 times BW, 1.8–2.2 times BH; BW 1.3–1.4 times BH; body depressed, ovoid with blunt snout and posterior edges of body in dorsal view; snout round in lateral view; eyes positioned and directed laterally; ED 0.6–0.8 mm, 35–53% of IOD; IOD less than IND; nares frontally positioned and directed laterodorsally; narial opening reniform in lateral view; END 0.8–0.9 mm. Spiracular tube sinistral, conical, projecting posterodorsally, its tip located 74–78 % of BL from snout. Lateral-line system conspicuous. Caudal musculature highest at body-tail junction, tapering posteriorly, terminating at tail tip; tail tip pointed; upper fin originating at junction of body and tail, gradually increasing in height to about 2/3 of tail; UTF 70–80 % of TMH; LTF 0.6–0.8 % of TMH; lower fin similar in proportion but slightly lower; MTH 16–21 % TL.
Mouth small, directed anteroventrally, oral disc width 0.66 mm (from edges of marginal papillae (0.38 from edges of sheaths). No labial teeth. Single posterior row of large and blunt marginal papillae with two pairs partially fused, one pair single and three lateral marginal papillae partially fused into a vertical fold on each side; anterior labium bare; submarginal papillae absent; jaw sheaths robust and finely serrated.
TABLE 5. Comparison of some diagnostic characters among species of the Dendropsophus parviceps group ( D. schubarti , D. microps , D. giesleri , D. timbeba were omitted given they
not cluster with the parviceps clade and are unambiguously morphologically distinct).
D. counani sp. nov. D. brevifrons D. bokermanni D. frosti D. koechlini D. ruschii D. luteoocellatus D. subocularis D. pauiniensis D. parviceps
Thighs dorsally dark grey with cream same as D. counani but large plain black plain black plain cream large large large large
colour blotches anterodorsally and generally less yellow/orange yellow/orange yellow/orange yellow/orange yellow/orange ventrally pale grey translucent conspicuous and with blotch on blotch on blotch on blotch on blotch on an orange spot anterior anterior surfaces anterior anterior ventral surface anteriorly in some surfaces of of thighs surfaces of surfaces of of tibia specimens thighs thighs thighs
Venter anteriorly white and granular and same as D. counani entirely cream entirely with black entirely entirely cream entirely cream granular with dark posterolaterally translucent grey and granular cream and flecks cream and and granular melanophores posteriorly
granular granular
Thighs smooth smooth smooth granular granular granular ??? ??? ??? smooth
texture
dark brown to grey, few scattered tan yellowish to dark light yellow to smooth finely smooth or smooth smooth tuberculate smooth tubercles brown, smooth mustard, shagreen with very
Dorsum smooth few nat
scattered
tubercles
pale grey on the axillary and the dorsolateral stripes in dorsolateral black flanks plain pale dorsolateral dorsolateral dorsolateral ??? brown with inguinal regions separated by a females stripes brown stripe in stripes stripes cream marks series of oblique dark and cream dorsally, most posteriorily
Flanks patches extending from the marbled specimens
dorsum, and no dorsolateral grey
stripes ventrally
19.40–21.73 M, 22.10–24.50 F 17.0–21.4 M, 20.1– 22.7 M, 25.6 F 21.1–25.2 17.5–23.8 24.5–27.9 23.1 M, 28.0 F 23.1 mm M, 20.2–20.3 M, 18.3 M, 26.1 23.2 F (Duellman & M, 25.9– M, 23.9– M, 26.7– 26.1 F 23.0–24.0 F mm F
) Crump 1974) 28.F 28.1 F 29.0 F
Canthal/ absent canthal and rostral canthal canthal rostral canthal and canthal canthal and canthal and absent
rostral rostral rostral rostral
Advertis Trills X = 0.30 s, 33–50 trills X = 0.47 s, 14 trills X = 0.32 ??? trills> 0.4s??? trills> 0.4s Trills> 0.4s??? Trills < 0.2 s ement notes/trill notes/trill (Duellman & s, 34–36
click series X = 0.45 s, 3–6 Crump 1974); 0.44 s, notes/trill
clicks, each with 1 to 3 notes 15–16 notes/trill clicks X =
click series X = 0.75 s, 0.62s, 5–7
4–5 single notes clicks single note
in QCAZ5848 clicks in
QCAZ26725
In life, tail musculature and the flanks of the body below a line extending from snout to the upper edge of fin are black with golden patches; back and dorsal side of the flanks golden with black flecks. The fin is translucent with golden fleck anteriorly and black patches on the posterior third; the iris is golden with a reddish transversal mark.
Tadpoles of Dendropsophus counani sp. nov. can be distinguished from those of D. koechlini , D. parviceps , D. ruschii , and D. brevifrons by the fusion of the lateral marginal papillae; from D. bokermanni by the absence of secondary papillae ventrally, and from D. subocularis by the fusion of the few ventral marginal papillae. They can also be distinguished from D. koechlini , D. parviceps , D. ruschii , D. bokermanni , and D. subocularis by the shape and the coloration of the fin (reaching its max height around 2/3rd of the length and harboring black pigmentation before the tip in D. counani ).
Distribution, ecology, and conservation. In addition to the material examined, above 13 additional occurrences in French Guiana were retrieved from the personal observations of various persons (Appendix 3), as well as three in Suriname ( Ouboter & Jairam 2012; pers. obs.), two in Amapá (acoustic record from J. Dias Lima; pers. obs.) and one in Guyana ( Cole et al. 2013) ( Fig. 1 View FIGURE 1 ). The absence of previous records in Amapá ( Dias Lima 2008) and Northern Pará ( Avila-Pires et al. 2010) is noteworthy. Lima et al. (2006) provide details on Dendropsophus brevifrons from Reserva Adolfo Ducke but the species probably belongs to the D. microcephalus group.. Our identification is based on the visual examination of the pictures, the video record provided, and personal examination by one of us (VGDO).
This species inhabits primary and secondary forest and is encountered only when breeding, in temporary ponds, as well as flooded parts of stream and river beds after heavy rain. Males call on vegetation (on small branches or leaves) overhanging or nearby water. They often start calling in the late afternoon and continue at night. Outside these reproduction activity events, the species likely dwells in the canopy and thus remains difficult to detect. Even when active, calling males often remain high in the trees and descend only after heavy rains. This is likely the cause for the failure to detect the species during previous surveys undertaken in Amapá ( Dias Lima 2008) and Northern Pará ( Ávila-Pires et al. 2010). Clutches (50– 79 eggs) are deposited on vegetation above water and tadpoles drop from the gelatinous clutch after a few days. Six days before hatching, clutches have already a typical elongated shape and exhibit drip-tips at the distal end ( Fig. 6 View FIGURE 6 d).
Dendropsophus counani View in CoL occurs from the Brazilian state of Amapá across French Guiana and Suriname to Central Guyana and is likely to extend its range into adjacent Brazilian states and eastern Venezuela. The single specimen from Belém (KU127846) that was examined by Duellman & Crump (1974) cannot be unambiguously identified as D. counani View in CoL sp. nov. This specimen, which was not included in the type series by Duellman and Crump (1974), was examined by us via pictures provided by the curators of the herpetological collection of KU and is very similar to specimens of D. counani View in CoL sp. nov. i.e. from the other side of the Amazon River. Nevertheless, some differences exist ( e.g. white flash mark on flanks and on dorsal side of legs are quite different). Given that there is only a single individual available from south of the Amazon River and molecular data is missing, it is difficult to draw any conclusions about the identity of the Belém population.
In French Guiana, Dendropsophus counani View in CoL sp. nov. occurs throughout the territory and can be locally abundant when breeding, as it is probably also the case in Suriname and Amapá. In Guyana, throughout the entire study period, the species appeared only sporadically (only on very few days/nights), very localized (only two localities), and in low densities (only 24 calling males during 393.5 hrs of standardized visual and acoustic transect sampling, or 0.06 individuals/transect hour) (Ernst et al. 2006; Ernst & Rödel 2008).
Both (B1) the extent of occurrence (> 20.000 km 2) and (B2) the area of occupancy (> 2000 km 2) of Dendropsophus counani View in CoL are largely over the threshold of the IUCN threatened categories. Moreover, its range lies in the Guiana Shield, which is relatively well preserved, the range of D. counani View in CoL is putatively continuous and no noticeable decline has been observed nor is expected in the upcoming years. Therefore, the species should be listed as Least Concern.
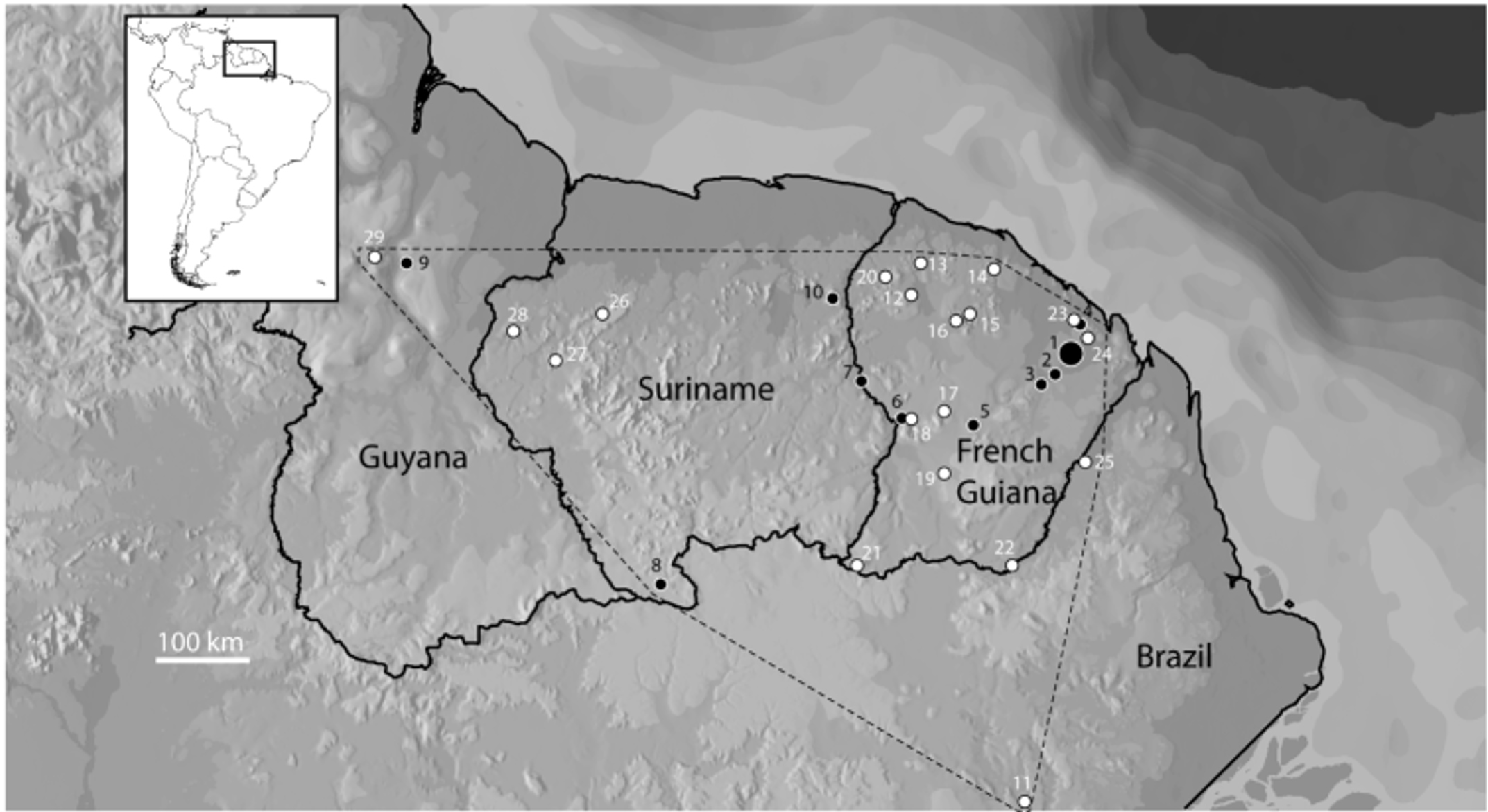





TABLE 3. Call variables obtained from four calls of Dendropsophus counani sp. nov. and calls of D. bokermanni and D. brevifrons.
| D. counani sp. nov. | D. bokermanni | D. brevifrons | |||
|---|---|---|---|---|---|
| Aratai Inini | Saül | Mabura | Ecuador | Ecuador | |
| Trill duration (s) | 0.348 (0.001) 0.310 (0.008) | 0.313 | 0.215 (0.001) | 0.326 (0.0275) | 0.443 (0.012) |
| Inter trill silence (s) | 6.635 (NA) 10.62 (8.866) | 23.9 (NA) | 1.593 (NA) | 52.553 | 0.975 (NA) |
| Trills/min | 10 (NA) 8 (NA) | 4 (NA) | NA | 1 (NA) | 3 (NA) |
| Notes/trill | 50 (0) 41.5 (1) | 44 (0) | 33 (0) | 34.0 (2.828) | 15.5 (0.5) |
| Note duration (s) | 0.0053 (0.001) 0.005 (0.000) | 0.005 (0) | 0.004 (0) | 0.005 (0) | 0.0113 (0) |
| Inter note silent (s) | 0.001 (0) 0.001 (0) | 0.001 (0) | 0.002 (0) | 0.003 (0) | 0.0177 (0) |
| Rate (notes/s) | 143.679 134.0 (2.549) | 140.4 | 153.1 (0.502) | 104.3 (2.202) | NA |
| Trill dom. freq. (Hz) | 3855 (2) 3960 (12) | 3578 (17) | 3915 (4) | 4204 (151) | 4243 (60) |
| Trill 2nd freq. (Hz) | 8038 (36) 7650 (102.) | NA | NA | 8167 (404) | 7990 (2) |
| Trill 3rd freq. (Hz) | 11922 (4) 11163 (726) | NA | NA | 12434 (33) | 12919 (428) |
| Click series duration (s) | 0.407 (0.004) 0.497 (0.140) | 0.539 | 0.331 (0.079) | 0.618 (0.072) | 0.751 (0.109) |
| Inter click series silence | 2.538 (0.585) 5.41 (0.532) | 2.192 (0.36) | 1.409 (0.312) | 2.807 (1.848) | 1.635 (0.262) |
| Click series/min | 24 (NA) 12 (NA) | 24 (NA) | 30 (NA) | 20 (NA) | 18 (0) |
| Clicks/series | 4 (0) 4.667 (1.528) | 4.5 (0.577) | 3.5 (0.577) | 6 (0.816) | 4.667 (0.471) |
| Notes/click | 2.062 (0.239) 2.622 (0.038) | 1.487 | 1.646 (0.105) | 1 (0) | 1 (0) |
| Inter click silence (s) | 0.103 (0.004) 0.114 (0.004) | 0.129 | 0.096 (0.006) | 0.110 (0.016) | 0.183 (0.009) |
| Rate (Clicks/s) | 9.835 (0.089) 9.355 (1.222) | 8.379 | 10.701 | NA | NA |
| Click series dom. freq. | 3796 (21) 3878 (83) | 3428 (46) | 3787 (3) | 4373 (48) | 4112 (18) |
| Click series 2nd freq. | NA 7801 (77) | NA | NA | 8598 (182) | 7970 (9) |
| Click series 3rd freq. (Hz) | 11506 (117) 10826 (NA) | NA | NA | 11634 (NA) | 12311 (4) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.