
Drosera maanyaa-gooljoo A.Fleischm. & T.Krueger, 2023
|
publication ID |
https://doi.org/10.11646/phytotaxa.618.1.3 |
|
DOI |
https://doi.org/10.5281/zenodo.8403482 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687D1-1E54-FFB9-FF42-532EFAF1EBAA |
|
treatment provided by |
Plazi |
|
scientific name |
Drosera maanyaa-gooljoo A.Fleischm. & T.Krueger |
| status |
sp. nov. |
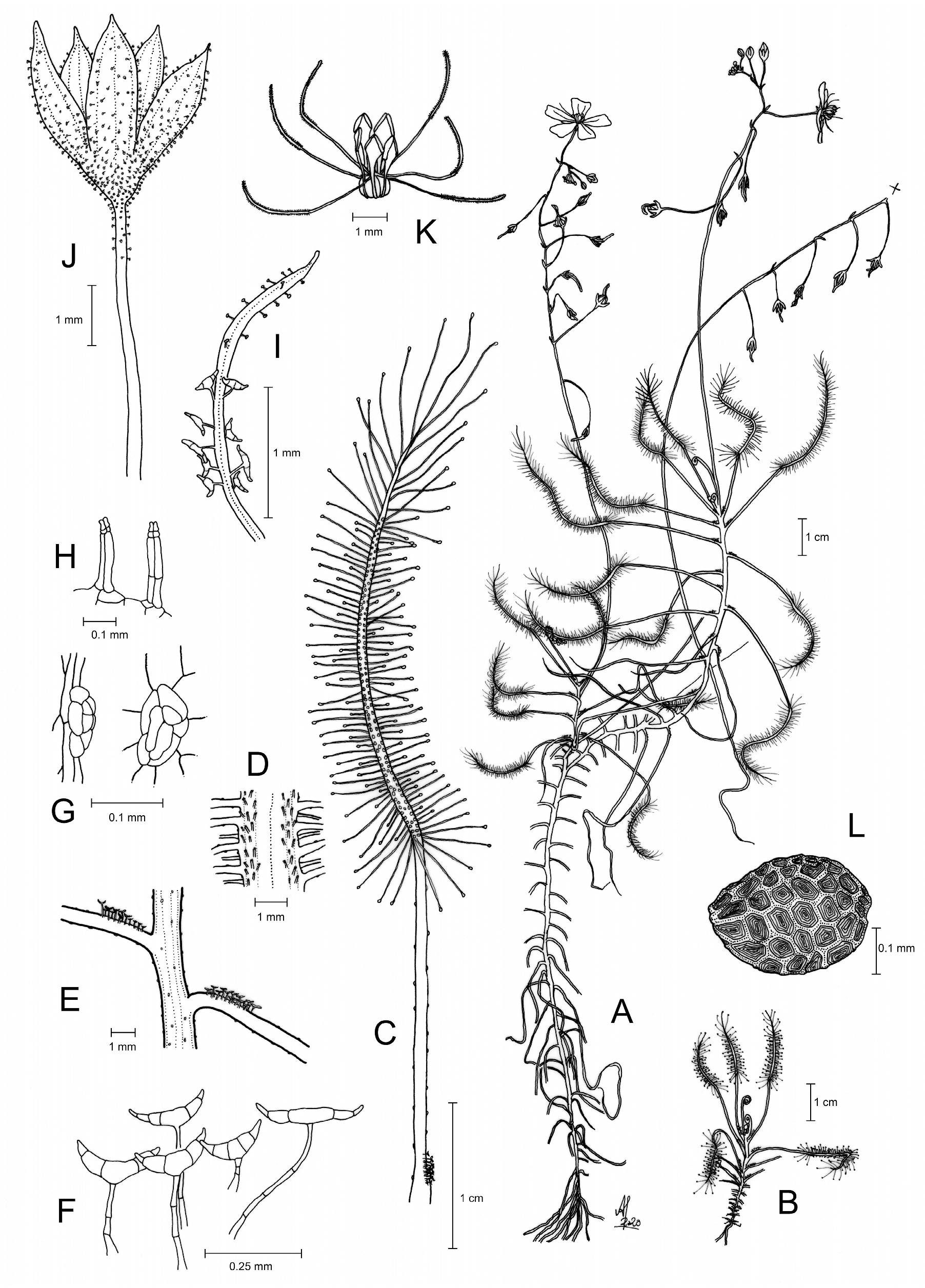
Drosera maanyaa-gooljoo A.Fleischm. & T.Krueger , sp. nov. ( Figs. 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ).
Type: — AUSTRALIA. Western Australia: Buccaneer Archipelago , [precise locality withheld for conservation purposes], within 10 m of stream, 18 June 1982, A.J.M. Hopkins BA 0248 (holotype PERTH 05615631 About PERTH !, isotype PERTH 09120807 About PERTH !) .
Diagnosis: — Drosera maanyaa-gooljoo differs from all other known members of Drosera section Arachnopus by (contrasting characters in parentheses): 1) its long-petiolate leaves with petioles exceeding or equalling the lamina length, hence making up about 50% or more of the total leaf length (petiole absent or reaching at most ca. 15–20% of the total leaf length); 2) a patch at the petiole base of stalked, multicellular, non-carnivorous trichomes with translucent, drying pale yellowish-brown, transversely crescent-shaped heads (the only other two species of D. section Arachnopus with similar patches of relatively large non-carnivorous trichomes or modified tentacles at their petiole bases are D. hartmeyerorum , with moriform heads, and D. barrettiorum with elliptic to hyperbolic-paraboloid heads); and 3) the lack of any obvious indumentum on stems, petioles, and peduncles except for scattered, microscopic, sessile glands with flattened hemispherical, multicellular gland head (stalked glandular trichomes of various shapes and eglandular trichomes found on stems, petioles, and peduncles of all remaining members).
Description: —Annual herb, 15–40 cm tall including inflorescence. Roots simple, short, slightly succulent, terete, poorly branched, one primary root and several lateral roots. Stem upright, self-supporting, leaning on nearby vegetation or becoming prostrate with age, up to at least 35 cm tall, simple or becoming branched from the middle in flowering individuals (four lateral branches, each floriferous, present in the holotype specimen), terete, 1.2–2.9 mm in diameter, yellowish green; stem, petioles and scape appearing glabrous by lacking any indumentum of stalked trichomes but covered with scattered sessile glands with flattened hemispherical, multicellular gland head (producing a hemispherical translucent secretion droplet in living individuals); internodes (0.1–)0.5–5.0(–8.0) mm long, much shorter in the lowermost 0.5–2.0 cm of stem. Leaves filiform, ca. 30–80 mm long in flowering-size specimens, straight but with an angle of ca. (10–)20–65(–80) degrees between petiole and lamina (angle often increasing with leaf age), vernation circinate; (10–)12–20(–26) active leaves present in flowering specimens; stipules absent; petiole linear, (14–)20–40(–45) mm long, 0.8–2.3 mm wide near base, gradually narrowing to 0.6–1.0 mm near lamina, green or yellowish green but adaxial base red (often also reddish near lamina), in section narrowly transversely triangular, 0.4–1.2 mm thick, abaxially convex, adaxial surface shallowly domed and with markedly raised midrib, senescent petioles persistent, patent; adaxial petiole surface glabrous except for a cluster of 20–38 stalked transversely crescent-shaped trichomes (“emergences”) at the petiole base; trichome stalk terete, red or translucent, slightly tapering towards the tip, (0.1–)0.2–0.6(–0.7) mm long (trichomes with shorter stalk towards the petiole base, the distal ones with longer stalk), 0.03–0.07 mm wide at its base, uniseriate; trichome head transversely crescent-shaped, translucent (turning pale yellowish-brown in dried material), 0.2–0.5 mm long, 0.08–0.15 mm wide, 3–5(–7)-celled, distal cells horn-like with pointed tip; petiole surfaces additionally covered with scattered sessile glands of the same type as present on stem; lamina narrowly linear-lanceolate with long acuminate tip, curved, (14–) 18–35 mm long (excluding tentacles), 0.6–1.5 mm at its greatest width (excluding tentacles), ca. 0.1 mm wide at the long-acuminate tip; lamina red (yellowish-green in freshly unfolded leaves, turning red with age), its adaxial surface covered with stalked, carnivorous, secretive capitate glands (tentacles), tentacle stalk red, gland head red, all tentacles with radially symmetric gland head, tentacles at lamina apex, base and lateral margins exceedingly long-stalked (longest towards tip and base of the lamina), 5–11 mm long, short-stalked tentacles on the inner lamina surface 0.1–0.3 mm long; lamina abaxial surface covered with two rows of biseriate double-tipped trichomes (type 5 pro parte s ensu Länger et al. 1995) ca. 0.1–0.3 mm long, with reddish stalk and translucent or yellowish tips. Inflorescences 1–4(7), each forming a many-flowered, bracteate scorpioid cyme ([13–]16–27 flowers per cyme); peduncle, rhachis and pedicels appearing glabrous, calyx and sepals glandular; scape 20–40 cm tall including peduncle, green to yellowish green; peduncle terete, 0.8–1.6 mm in diameter, ascending vertically (± parallel to the stem axis even in old inflorescences), glabrous except for the sessile glands; fruiting pedicels spaced by 2–18(–37) mm; pedicels terete, base often concaulescent, (7–)9–18(–23) mm long at anthesis and in fruit, 0.3–0.6 mm in diameter, covered with scattered sessile glands of the same type as on stem and petioles and additionally with few stalked capitate trichomes towards the calyx (corresponding to those found on sepals); pedicels straight at anthesis becoming more or less arcuated or reflexed in fruit (arcuated horizontal/slightly downwards from rhachis, then sharply curving upright near the seed capsule; pedicels of unfertilised flowers pendulous). Bracts very narrowly oblanceolate, lateral margins entire, apex obtuse or acute and irregularly crenate, (1.2–) 1.5–2.7 mm long, 0.2–0.5 mm wide, adaxial surface glabrous but basally with 5–15 stalked transversely crescent-shaped trichomes (corresponding to those found at the petiole base; lowermost bracts on a scape occasionally lacking such trichomes), abaxial surface glabrous in the lower 2/3, upper 1/3 covered with capitate trichomes (corresponding to those found on the sepals); every pedicel except that of the lowermost flower subtended by a bract and bracts persistent in fruit. Flowers 11–15 mm in diameter. Sepals 5, lanceolate to elliptic, margins entire, with acute (rarely obtuse or acuminate) apex, 2.1–3.5 mm long, (0.5–) 0.8–1.3 mm at their greatest width, abaxial surface densely covered with stalked capitate trichomes ca. 0.1–0.2 mm long with red (or rarely translucent), uniseriate stalk and translucent-white, spherical gland head ca. 0.05 mm in diameter; sepals basally adnate to form a flat-campanulate calyx. Petals 5, obovate to broadly obovate (sometimes cuneate), apical margin truncate or emarginate and finely crenulate, 5.2–7.5 mm long, 3.0– 4.2 mm wide at their widest point, pink or pale pink; petal aestivation imbricate, contorted. Stamens 5, alternipetal, oblanceolate to spathulate, 2.0– 2.5 mm long, 0.7–1.0 mm wide; filaments very narrowly cuneate, 1.1–1.5 mm long, 0.2–0.3 mm wide, white; anthers bithecate, 0.8– 1.2 mm long, connective dilated, triangular, 0.6–0.8 mm wide, white, thecae pale yellow; pollen pale yellow. Ovary 3- carpellate, fused, subglobose, shallowly 3-lobate in outline, 1.0– 1.2 mm in diameter, glabrous, yellowish green. Styles 3, each basally divided into 2 entire style-arms; style-arms flagelliform, terete, 1.7–2.2 mm long (excluding stigmatic portion), 0.08–0.12 mm in diameter, translucent white, glabrous, positioned horizontally or arcuately curved upwards, graduating into stigmatic part; stigmas 0.9–1.8 mm long, 0.08–0.20 mm in diameter, translucent white, papilliate. Seeds numerous, 300–380(–410) µm long, 250–280 µm wide, broadly ellipsoidal, testa black with slightly iridescent shine of the periclinal walls, testa surface isodiametrically reticulate, with anticlines thin and pale brown, periclinal walls tabular, microscopically rugose.
Etymology: —The specific epithet “ maanyaa-gooljoo ” was selected by the Dambimangari and Mayala Traditional Owners. The word “ maanyaa ” means centipede in the Worrora language, and is the name given to this species by the Dambimangari Traditional Owners. It references the lamina of the species, which is covered with exceptionally long tentacles at the lamina base and tip, thus resembling certain long-legged centipedes (e.g., of Scutigeridae ). The second word of the epithet, “ gooljoo ”, was selected by the Mayala Traditional Owners and means grass in the Bardi language. Both words of the epithet stand independently, and thus the use of a hyphen is permitted in this case according to ICN rule 60.11 ( Turland et al. 2018; Turland pers. comm. 2023).
Taxonomic notes: — Drosera maanyaa-gooljoo clearly belongs in D. section Arachnopus , as is evident from its overall habit of narrowly linear-lanceolate, exstipulate leaves spaced along elongated stems, as well as from floral morphology with six entire, long, flagelliform style-arms ( Planchon 1848, Lowrie 2014) and the presence of biseriate double-tipped trichomes. However, the exact affinity to known species of that section is puzzling based on morphology alone. Drosera maanyaa-gooljoo is unique in several characters, such as (1) the long-petiolate leaves with an angulate transition between petiole and lamina, (2) very long marginal tentacles not only present towards the leaf tip (as realised in all other members of the affinity) but also present at the lamina base, and (3) lacking any obvious indumentum of stalked trichomes on stems, petioles and peduncles, which are instead covered by (4) unique, sessile glands with multicellular secretive gland head. These four characters are not paralleled in any other known species of D. section Arachnopus .
The presence of clusters of distinctive, multicellular, translucent- or yellow-headed “emergences” at the petiole base (and at the bases of the bracts along the scape) is also found in Drosera barrettiorum and D. hartmeyerorum , yet each species exhibits a very different, unique “emergence” type ( Fig. 6 View FIGURE 6 ; Table 1 View TABLE 1 ). While the “emergences” in D. barrettiorum and D. hartmeyerorum are clearly homologous and represent modified tentacles (their stalks being vascularised), the stalked transversely crescent-shaped trichomes of D. maanyaa-gooljoo are non-vascularised. While much larger in size, these trichomes of D. maanyaa-gooljoo also share morphological similarities with the T- or Yshaped trichomes found in D. serpens Planchon (1848: 204–205) ( Fig. 6 View FIGURE 6 ) and D. finlaysoniana Wallich (1831 : n. 3752) ex Arnott in Hooker (1837: 314) ( Krueger & Fleischmann 2021). Seed shape, size, and ornamentation of D. maanyaa-gooljoo is similar to D. barrettiorum and D. hartmeyerorum : these three taxa share very small black seeds with regular isodiametric testa reticulation and thin, pale brown, shallowly raised anticlinal walls ( Fig. 7 View FIGURE 7 ), while all remaining species of D. section Arachnopus exhibit a different seed morphology and seed size ( Susandarini et al. 2002, Barrett & Lowrie 2013, Lowrie 2014, Krueger & Fleischmann 2021; Fig. 7 View FIGURE 7 ). However, only D. barrettiorum has an iridescent shine of the testa periclinal walls like D. maanyaa-gooljoo , while the seeds of D. hartmeyerorum are dull black ( Fig. 7 View FIGURE 7 ). Drosera aurantiaca Lowrie (2014: 1268) also produces seeds with isodiametrically reticulate testa with iridescent shine of the periclinal walls, yet it has slightly larger seeds than the three above-mentioned species ( Fig. 7 View FIGURE 7 ), and lacks any clusters of conspicuous trichomes or modified tentacles on its leaves.
However, Drosera maanyaa-gooljoo displays very long petioles (the longest and most distinctly developed in any member of D. section Arachnopus ) that exceed or equal the lamina length in juvenile as well as in flowering-size plants. This is strongly contrasting D. barrettiorum and D. hartmeyerorum whose petioles are absent (i.e., the tentaclebearing part of the leaf reaches all the way to the stem; Fig. 6 View FIGURE 6 ; Lowrie 2014). In this respect, D. maanyaa-gooljoo more closely resembles well-petiolate species like D. serpens . Interestingly, seedlings and juvenile plantlets of all species of D. section Arachnopus have petiolate leaves, even those which develop sessile laminae when reaching flowering size, e.g., D. finlaysoniana (A. Fleischmann & T. Krueger pers. obs.). Hence, petiolate leaves can most likely be considered symplesiomorphic in the section. However, in all previously known species with petiolate leaves, the petiole length only reaches a small fraction of the total leaf length (ca. 5–20%); the petiole in D. maanyaa-gooljoo exceeds or equals lamina length and thus comprises ≥ 50% of the total leaf length.
Another unique feature of Drosera maanyaa-gooljoo is the lack of any indumentum of stalked trichomes on stems, petioles, and scapes which thus appear fully glabrous. Only the sepals, bracts, and the uppermost part of the pedicels bear stalked capitate trichomes ( Fig. 3F,G,L View FIGURE 3 ). This strongly contrasts with all remaining members of D. section Arachnopus , which have stems, petioles, and scapes covered with various types of indumentum of eglandular and glandular trichomes, often with species-specific patterns and trichome types ( Barrett & Lowrie 2013, Lowrie 2014, Lowrie et al. 2017, Schlauer et al. 2018, Krueger & Fleischmann 2021). The only other species from that affinity which likewise has an entirely glabrous peduncle is D. glabriscapa Lowrie (2014: 1271) . However, D. glabriscapa bears a dense indumentum of short hispid, conical, eglandular trichomes on its stems and petioles ( Lowrie 2014).
Based on common seed characteristics, we hypothesise a close affinity of the four species Drosera aurantiaca , D. barrettiorum , D. hartmeyerorum and D. maanyaa-gooljoo . Additionally, all but D. aurantiaca are further united by the presence of conspicuous clusters of translucent- or yellow-headed “emergences” with species-specific shape of the gland head at the base of their petioles and bracts. A close phylogenetic affinity of D. aurantiaca , D. barrettiorum and D. hartmeyerorum is further corroborated by their common phytochemistry (presence of methyljuglone as the sole naphthoquinone compound; Schlauer et al. 2018, Schlauer & Fleischmann 2022).
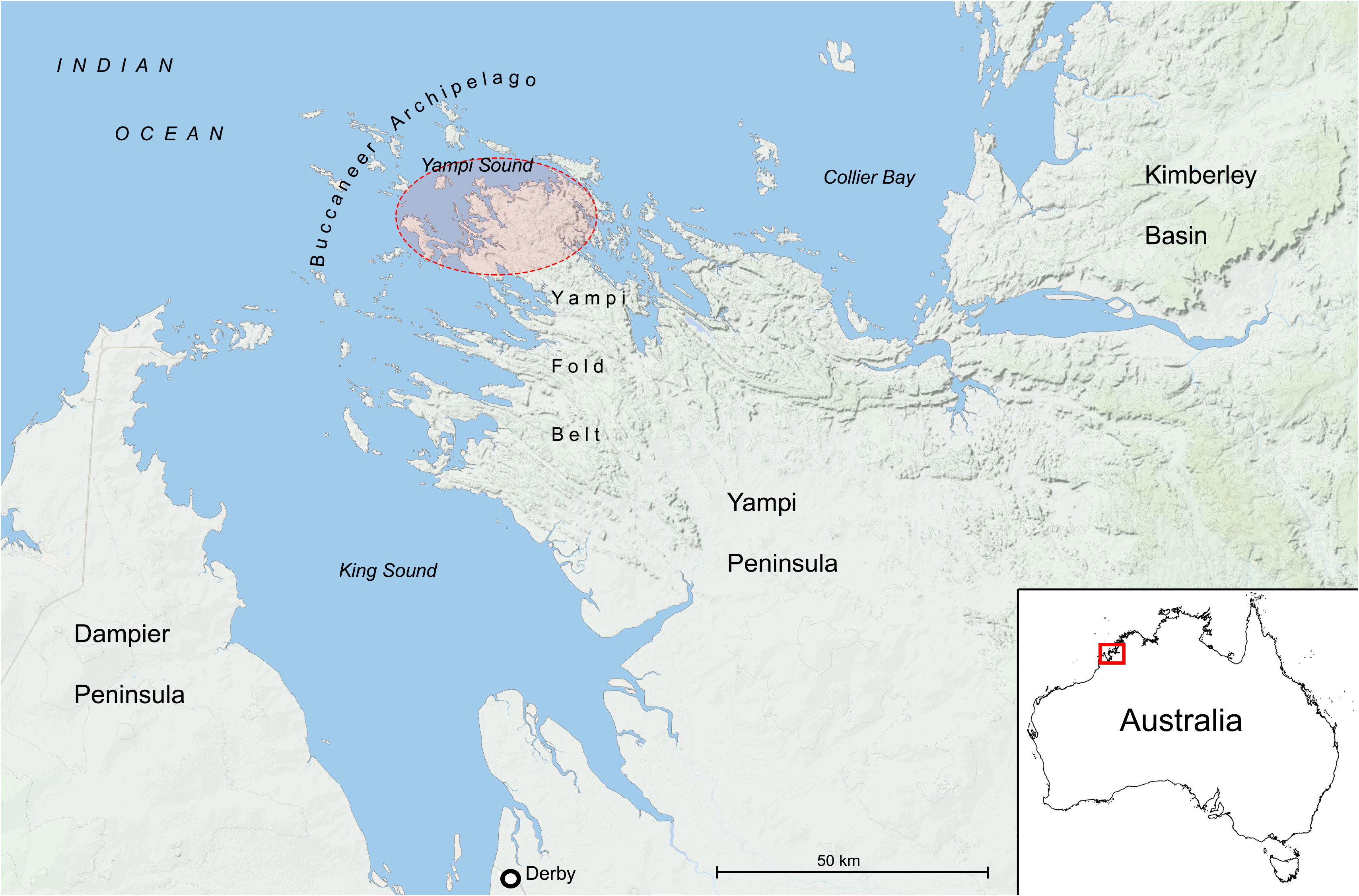
Distribution and habitat: —Endemic to the Kimberley Region of tropical northern Western Australia. Drosera maanyaa-gooljoo is known from only two localities ca. 20 km apart: one from an island in the Buccaneer Archipelago, and one from the northern Yampi Peninsula on the Australian mainland. Thus, the species is only known from areas located adjacent to the Yampi Sound of the Indian Ocean ( Fig. 5 View FIGURE 5 ). The species is most likely endemic to the northern Yampi Peninsula and the Buccaneer Archipelago, which are geologically and edaphically distinct from the neighbouring Dampier Peninsula and the Kimberley Plateau ( Tyler & Griffin 1992, Wilson 2013).
At the northern Yampi Peninsula locality, Drosera maanyaa-gooljoo occurs in wet areas alongside and within the bed of a shallow sandstone creek in shallow sandy soil over sandstone. Accompanying plants denoted include Grevillea spp. ( Proteaceae Juss. ), Corymbia polycarpa (F.Muell.) K.D.Hill & L.A.S.Johnson , Eucalyptus brachyandra F.Muell. , Melaleuca sp. ( Myrtaceae Juss. ), Templetonia hookeri Benth. ( Fabaceae Lindl. ), Owenia vernicosa F.Muell. ( Meliaceae Juss. ), and Distichostemon hispidulus Baill. ( Sapindaceae Juss. ). The habitat of the type population in the Buccaneer Archipelago is assumed to be very similar, described on the specimen label as “within 10 m of [a] stream”. At the northern Yampi Peninsula site, D. maanyaa-gooljoo was observed growing syntopically with the carnivorous plants Drosera burmannii Vahl (1794: 50) and Utricularia caerulea L. ( Lentibulariaceae Rich. ).
Other species of Drosera section Arachnopus recorded from Yampi Peninsula include D. barrettiorum , D. margaritacea Krueger & Fleischmann (2021: 57) , and D. serpens s.l. ( Western Australian Herbarium 1998 +, Lowrie 2001 [all taxa from D. section Arachnopus were denoted as “ D. indica ” at the time], Krueger & Fleischmann 2021). Of these, only D. barrettiorum is known to inhabit similar periodically flooded habitats near or within sandstone creek beds.
Ecology and phenology: —Annual, like all species from Drosera section Arachnopus . Flowering has been recorded in June and July, but likely extends from May until at least September. The species likely germinates following declining water levels in its creek habitat towards the end of the wet season in March or April, with individuals probably continuing to grow and flower until either the habitat dries out or until the onset of seasonal wet season flooding (typically in December). In June 2022, adult plants in flower and fruit were observed alongside newly-germinated seedlings ( Fig. 2 View FIGURE 2 ); thus, this species may continuously regrow from seed if persistent wet to moist soil conditions occur. Similar observations have been made for other members of D. section Arachnopus , including D. indica ( Ruan 1991) and D. finlaysoniana ( Krueger & Fleischmann 2021) .
Conservation status: —Listed as Priority One (poorly-known species) under Conservation Codes for Western Australian Flora ( Western Australian Herbarium 1998 +), under the phrase-name “ Drosera sp. Yampi Peninsula (A.J.M. Hopkins BA 0248)”. Data deficient (DD) following IUCN (2012), however, under possible category of threat because of small population sizes, and in need of further survey. Drosera maanyaa-gooljoo is currently only known from two localities, separated by ca. 20 km, both situated on exclusive possession Native Title lands belonging to Aboriginal peoples. Permission must be given by the Traditional Owners before the area can be accessed, and an Aboriginal Affairs Planning Authority (AAPA) permit must additionally be obtained for travel to the northern Yampi Peninsula. The type locality in the Buccaneer Archipelago is situated in the Mayala Traditional Owners’ exclusive Native Title determination. The second locality in the northern Yampi Peninsula is located within the Wotjalum Part III Aboriginal Reserve and part of the Dambimangari Indigenous Protected Area (IPA) which is managed by the Dambimangari Traditional Owners and Dambimangari Rangers. Only the latter population has been recently confirmed, with ca. 300 plants observed by the authors in June 2022. Population size at the type locality is unknown. It is noteworthy that recent floristic surveys of the area failed to locate this distinctive taxon ( Keighery et al. 1995, Barrett et al. 2001, Lowrie 2001, Lyons et al. 2014). Furthermore, a close examination of other Drosera specimens from the area in PERTH, as well as at national and international herbaria, did not identify any additional collections of this taxon. No populations occur on land managed by the Western Australian Department of Biodiversity, Conservation and Attractions (DBCA), and Drosera maanyaa-gooljoo (under the phrase-name “ Drosera sp. Yampi Peninsula”) has thus been listed as a possibly threatened species in urgent need of further survey effort (Priority One: Poorly-known species; Western Australian Herbarium 1998 +). Potential threats to this species may include the projected increase in the magnitude of extreme flooding events ( Moise et al. 2015) given the species’ preferred creek bed habitat, but also inappropriate fire regimes. Illegal collection may pose an additional threat in the future given the small reported population size and the unfortunate propensity for such carnivorous plant species to be rapidly targeted by poachers to provide plants for the horticultural market ( Cross et al. 2020).
Additional specimens examined: — Drosera maanyaa-gooljoo (paratypes):— AUSTRALIA. Western Australia: [northern Yampi Peninsula, precise locality withheld for conservation purposes], 02 August 1994, C. Done 847 ( PERTH 03783715 About PERTH !) [three juvenile individuals, sterile]; northern Yampi Peninsula [precise locality withheld for conservation purposes], 09 June 2022, T. Krueger & A. Fleischmann 51 ( PERTH 09546367 About PERTH !) [one fertile individual] .





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |