
Pediobius Walker, 1846
|
publication ID |
https://doi.org/10.11646/zootaxa.4438.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:688C9DA1-BE32-4FC6-B726-65CA36358473 |
|
DOI |
https://doi.org/10.5281/zenodo.5959787 |
|
persistent identifier |
https://treatment.plazi.org/id/03F687C9-FA3E-B467-FF27-68B0120FC934 |
|
treatment provided by |
Plazi |
|
scientific name |
Pediobius Walker, 1846 |
| status |
|
Pediobius Walker, 1846 View in CoL View at ENA
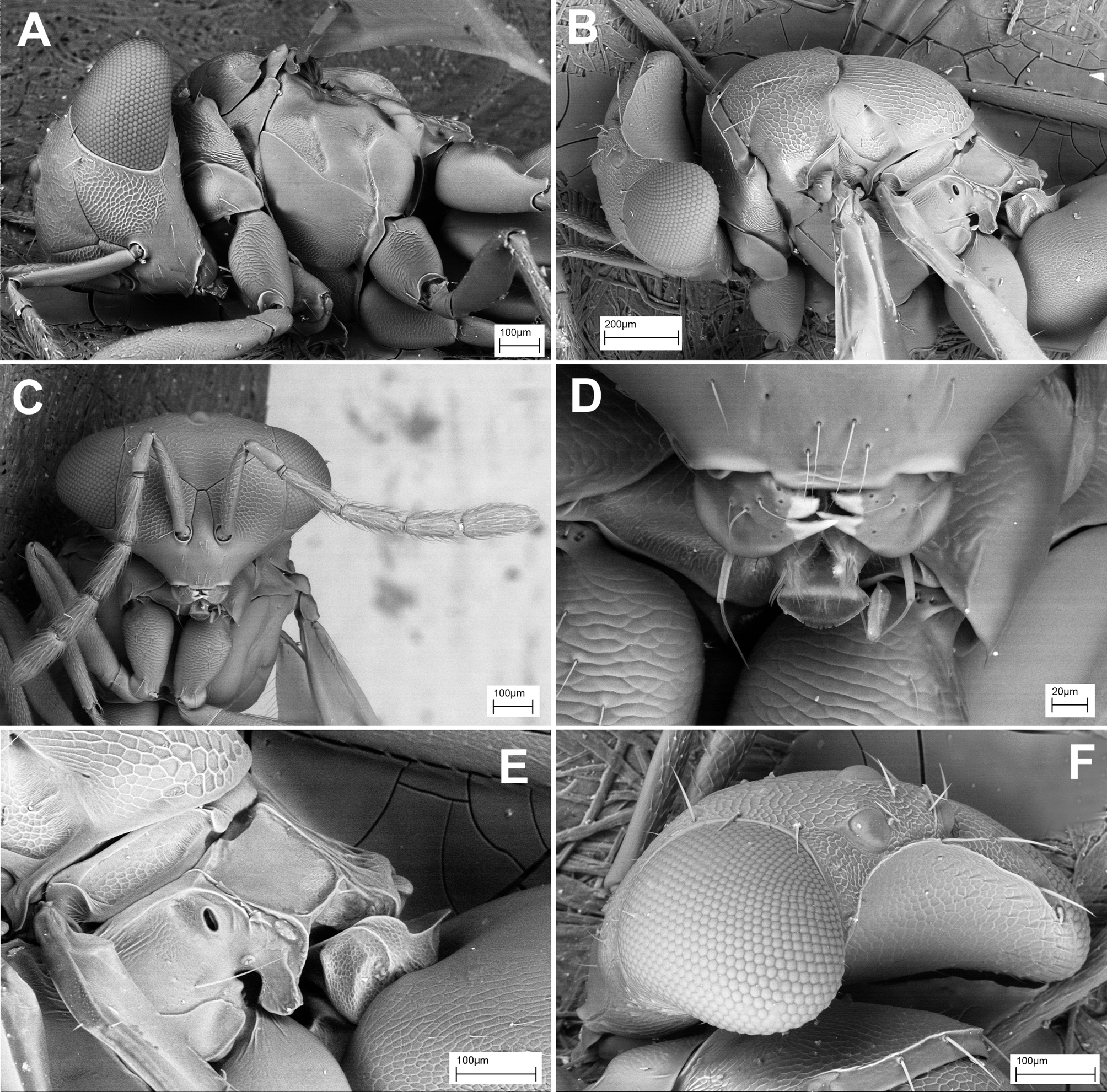
( Fig. 2 View FIGURE 2 )
Microterus SPinola, 1811: 151 –152. TyPe sPecies: Diplolepis petiolatus SPinola, by subsequent designation of Gahan & Fagan 1923: 89. Synonymized by Bouček 1965: 7. Regarded as nomen oblitum in favor of Pediobius walker by Hansson 2002: 146.
Pediobius walker, 1846 : 184 (as a subgenus of Entedon Dalman ). TyPe sPecies: Entedon ( Pediobius) imbreus walker, by subsequent designation of Ashmead 1904: 344, 384.
Pleurotropis Förster, 1856: 78 , 82. TyPe sPecies: Pleurotropis isomerus Förster View in CoL , by subsequent designation of Förster 1861: xxxvii. Synonymized by Ferrière 1953: 400.
Pediobomyia Girault, 1913: 155 View in CoL . TyPe sPecies (by original designation): Pediobomyia darwini Girault. Syn. View in CoL n.
Rhynchentedon Girault, 1919: 166 View in CoL . TyPe sPecies (by original designation): Rhynchentedon maximum Girault. Syn. n.
For full synonymy and diagnoses see Hansson (2002), Hansson & Nishida (2002), Cao et al. (2016).
Pediobius is rather species-rich and morphologically diverse, and the species described below expand the concepts of morphological diversity within the genus. If all data is combined, the genus may be diagnosed as follows ( Fig. 2 View FIGURE 2 ): body dark, black or with metallic tint ( Fig. 2A View FIGURE 2 ); head with complete frontal sulcus, more or less flat (most species, Fig. 2D View FIGURE 2 ) or largely smooth, convex ( afroteres group, Figs 9B–D View FIGURE 9 , 10A, C, D View FIGURE 10 ); lower face slightly (most species, Fig. 2D, F View FIGURE 2 ) or considerably narrowed (groups askari : Figs 12C View FIGURE 12 , 13A, C, D View FIGURE 13 , 14E, F View FIGURE 14 ; afroteres : Figs 9B–D View FIGURE 9 , 10A, C, D View FIGURE 10 ; maximus : Fig. 3B View FIGURE 3 , darwini : Fig. 3D, E View FIGURE 3 ); pronotal collar marked by a weak or strong carina ( Fig. 2B View FIGURE 2 ); propleurae widely curved over lateral panel of pronotum ( Fig. 2B View FIGURE 2 , pl1/f); mesoscutum without a median groove, with notauli marked at least by depressions (often traceable as deep marginated foveae); mesoscutellum mostly without median groove, lightly or coarsely reticulated, occasionally with lateral groves ( P. alcaeus Walker , P. bisulcatus Cao & Zhu ); mesosternum with narrowly extended anterior border slightly overlapping posterior border of prepectus ( Fig. 2B View FIGURE 2 , eps2/p); transepimeral sulcus present, grooved, weakly or strongly curved ( Fig. 2B View FIGURE 2 , tps); metapleuron with a tooth-like protrusion; propodeum with lateral plicae and small anterior foveae ( Figs 2C, E View FIGURE 2 , 18 View FIGURE 18 ); median area of propodeum with two carinae diverging posteriorly (most species, Fig. 2C, E View FIGURE 2 ), which may be modified as a raised keel ( alcaeus species group) or narrow submedian strip ( darwini group, see below); posterior part of propodeum elongate in varying degrees and often forming a delimited area, ‘nucha’ ( Figs 2C, E View FIGURE 2 , 3C View FIGURE 3 ); metasomal petiole distinct, robust and sculptured, often with raised upper edges ( Figs 2C, E View FIGURE 2 , 3C View FIGURE 3 , 18 View FIGURE 18 ).
Hosts and biology. The species of the genus are characterized by conceivably the widest host spectrum among Entedoninae : the reliable records range from larvae and pupae of Lepidoptera, Diptera and Coleoptera to larvae of thrips and egg sacks of spiders; hyperparasitoids are also common among species of the genus (Bouček 1965; Noyes 2017). Ironically, the data on immature stages are limited for such a biologically diverse genus. The available records suggest that Pediobius species are endoparasitoids with three larval instars (e.g., Bledsoe et al. 1983).
Revisions. The first comparative revision of Pediobius was provided by Bouček (1965) for its European species; which is still mostly up to date half a century after its publication, apart from a few nomenclatural changes. The Nearctic species were reviewed by Peck (1985) and the species occurring in the Japanese islands were keyed by Kamijo (1986). These reviews remain the main inventories for these regions. The Australasian species were listed and reviewed by Bouček (1988), and later reviewed by C. Burwell in his PhD thesis (Burwell 1995, unpubl.). The most recent comparative reviews are the revisions of Neotropical species by Hansson (2002) and of the Chinese species by Cao et al. (2017). The species from Indomalayan ( Oriental) realm were reviewed by Kerrich (1973) in his revision of the tropical and subtropical species of this genus. The Afrotropical species were very selectively reviewed and many hitherto described species as well as major part of museum materials were omitted in that same article. So, the Afrotropical fauna remains one of the most challenging parts of the species diversity of the genus.
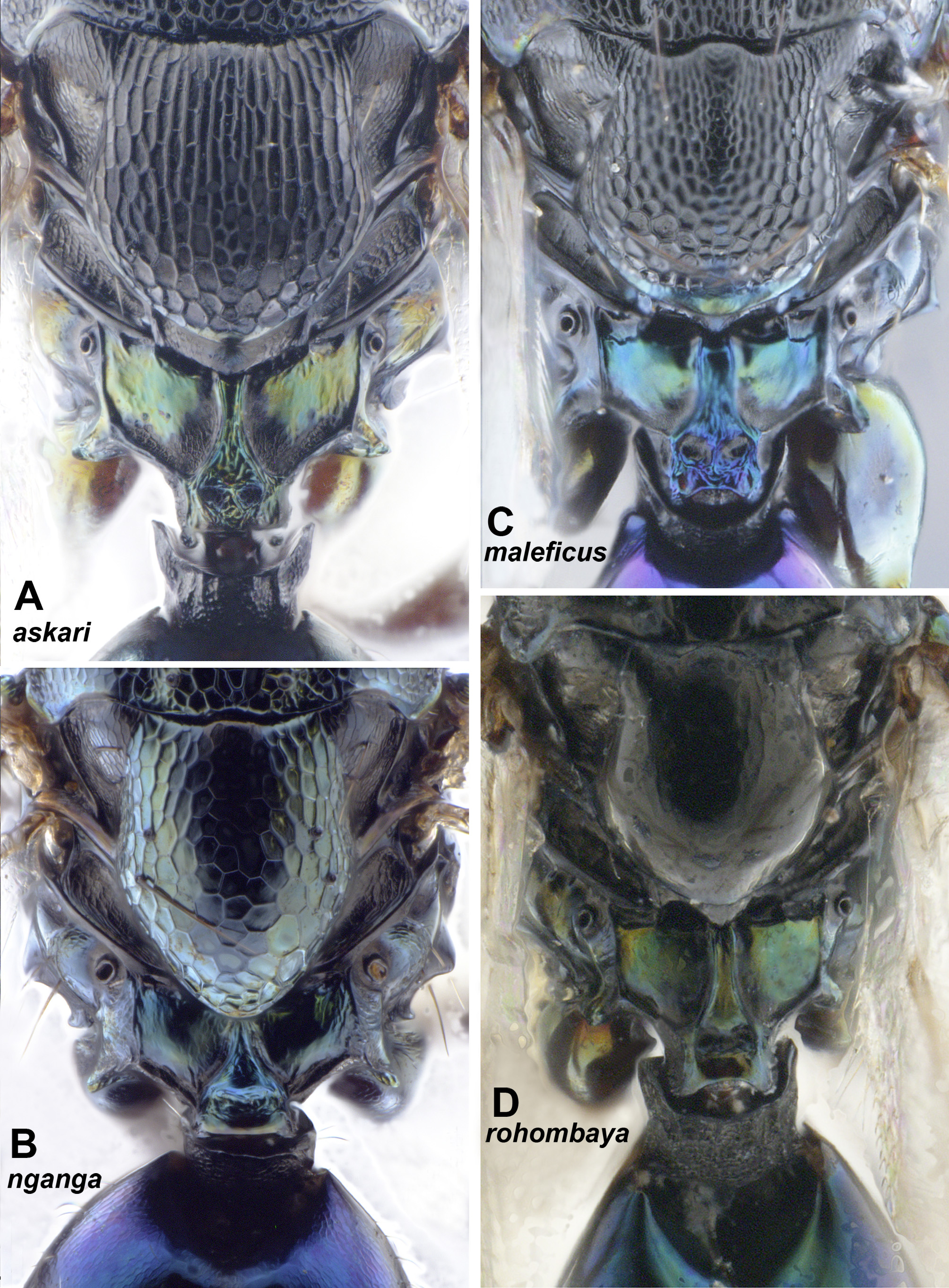
Comparative remarks. Pediobius Walker is the core genus of its genus complex, which was proposed to include also Rhynchentedon Girault , Pediobomyia Girault , Myrmokata Bouček and Microdonophagus Schauff (Gumovsky 2001) . The complex was characterized (Gumovsky 2001, 2003) by the wide and robust propleural flange (bending to anterior margin of the pronotal panel, Fig. 2B View FIGURE 2 , pl1/f), the anterior edge of mesosternum being narrowly extended and overlapping the prepectus ( Fig. 2B View FIGURE 2 , eps2/p), the lateral metapleural callus toothed, the metasomal petiole wide, robust, the propodeum with submedian foveae and more or less developed submedian propodeal carinae ( Figs 2C, E View FIGURE 2 , 3C View FIGURE 3 , 4E View FIGURE 4 ). Hansson et al. (2011) suggested sister-group relationships between the genera Horismenus and Microdonophagus , which were supported by the sculpture of propodeum in the newly described species of the latter. The close relationships between Pediobius , Rhynchentedon and Pediobomyia were confirmed by Burks et al. (2011); however, the hierarchy of these relationships was not established due to disagreement of the phylogenetic patterns resultant from different analyses. Rhynchentedon , Pediobomyia and Myrmokata were also characterized by the narrowed lower face resembling the species of Pediobius described below. The wide range of the head shape and sculpture, as well as the variety of other characters (e.g. the sculpture of dorsum) in the species described below in the marjoriae , afroteres and askari groups, suggest reconsideration of the status of some genera of the Pediobius complex.
The genus Rhynchentedon was described by Girault (1919) and then reviewed by Bouček (1988) and Gumovsky (2001, 2004). The genus is characterized by the narrowed and elongate lower face with concave genae, the reduced mandibles, wide propleural flange, densely setose axilla, transepimeral sulcus continued into a dropshaped fovea, and the propodeum with spiracles situated in depressions and with diverging submedian carinae ( Fig. 3A–C View FIGURE 3 ; Gumovsky 2001). In general, apart from the narrowed lower face and reduced mandibles, Rhynchentedon species resemble coarser sculptured and more setose representatives of Pediobius . The concave genae covered with dense setae may be considered a distinctive feature of Rhynchentedon ( Fig. 3B View FIGURE 3 ). However, the presence of the very weakly setose genae in R. narendrani Gumovsky hampers the diagnostic value of this character (Gumovsky 2004). The excessive setation, which is present in places in R. maximus Girault and R. achterbergi Gumovsky , is shared by some Afrotropical Pediobius species, namely P. setigerus Kerrich and P. multisetis Bouček. The reduced mandibles lacking the cutting edge, are also present in some species of Pediobius , e.g. P. derroni Bouček ( Fig. 2F View FIGURE 2 , md). Finally, different degrees of elongation of the face occur in species of Pediobius , as described below ( Fig. 17A–D View FIGURE 17 ).
The genus Pediobomyia was erected by Girault (1913) for his species P. darwini Girault described from Australia. The genus was briefly discussed by Bouček (1988) and Gumovsky (2001). Hansson (2002) updated its diagnosis for two newly described Neotropical species. Narendran et al. (2007) reviewed this genus based on materials from the Indomalayan (Oriental) realm and synonymized Bomyiabius Narendran (described earlier: Narendran 2004) with Pediobomyia . If all concepts are combined, the genus may be diagnosed as a group of entedonines with the lower face narrowed ventrally towards a very small mouth opening, mandibles lacking the cutting edges, scrobal and frontal sulci short grooved ( Fig. 3D, E View FIGURE 3 ), metascutellum with longitudinal carina at lateral margins, propodeum elongate with plicae converging towards petiolar foramen, submedian propodeal carinae diverging posteriorly or subparallel, and propodeal spiracle placed on an elevation, with long pliciform protrusion below it (Bouček 1988, Gumovsky 2001, Hansson 2002). Also, Hansson (2002) suggested that the pronotum is strongly reduced and hardly visible in dorsal view in the Neotropical species assigned to Pediobomyia . The genus has been considered as morphologically distinct, if its species were compared to the described species of Pediobius or Rhynchentedon (Gumovsky 2001) . However, if the representatives of Pediobomyia are compared to the species of afroteres group of Pediobius (described below: Figs 8–11 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 ), the differences in head shape and propodeum appear gradual. Furthermore, the representatives of the marjoriae ( Figs 5–7 View FIGURE 5 View FIGURE 6 View FIGURE 7 ) and askari ( Figs 12–15 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 ) groups which are also described below, demonstrate a range of character states being gradual between the afroteres group and the most other Pediobius species.
A direct comparison of the species of Rhynchentedon and Pediobomyia with the species of Pediobius mentioned above suggests the gradual nature of the characters used to distinguish them. So, notwithstanding their odd general appearance, the species which have been classified in Rhynchentedon and Pediobomyia (as well as the species described below) appear derived groups within speciose and diverse genus Pediobius . Therefore, morphological evidence suggests that keeping all these species in separate genera putatively renders Pediobius paraphyletic.
The close relationships of Pediobius with Rhynchentedon and Pediobomyia were demonstrated by Burks et al. (2011) using molecular markers. The genera were always closely related in all phylogenetic analyses provided by the authors, and at least in the Bayesian molecular-only results and in the combined morphological and molecular results, Rhynchentedon + Pediobomyia appeared as a derived node within Pediobius with high bootstrap support (Burks et al. 2011). So, based on original morphological data and in part on results of molecular research of other authors, the generic names Rhynchentedon Girault and Pediobomyia Girault are hereby considered junior synonyms of Pediobius Walker ( syn. n.). The following new combinations are resultant from this synonymy: P. maximus (Girault) , P. achterbergi (Gumovsky) , P. narendrani (Gumovsky) , P. brevicaulis (Hansson) , P. canaliculatus (Hansson) and P. darwini (Girault) ( comb. n.).
Bomyiabius frontus Narendran , Pediobomyia budaicus Narendran and Pediobomyia lankicus Narendran , described from Indomalayan realm by Narendran (2004) and Narendran et al. (2007), were reported as differing from Pediobomyia darwini Girault in rather superficial or misinterpreted characters. For instance, P. darwini was reported as differing from three other species listed above in having F2 shorter than pedicel and darker legs with metallic tint, and the three species differ chiefly by the relative length of F1. However, these characters appear variable within series collected in various regions, their dimensions (relative length of F1 and F2) depend on measurement points (including or excluding petiole of the funiculars) and the degree of collapse when specimens are dried. Based on available descriptions and the morphology of the studied materials from Australasian, Indomalayan and Afrotropical realms (partly listed in Gumovsky 2001), all three specific names mentioned above are proposed to be junior synonyms of Pediobius darwini (Girault) (in its new combination, Fig. 3D, E View FIGURE 3 ) ( syn. n.). The species that have been previously classified in Rhynchentedon and Pediobomyia are proposed to represent two species groups within Pediobius : maximus and darwini , respectively.
Myrmokata Bouček was described as an aberrant ant-associated Afrotropical entedonine genus for M. diparoides Bouček from Cameroon (Bouček 1972). The material of M. diparoides from RMCA ( ♀, “Coll. Mus. Congo, Basoko, Yawinawina, VIII.1948, P.I.G. Benoit”, Fig. 4 View FIGURE 4 ) expands distribution range of this species to DRC ( new record). Myrmokata was characterized by the considerable reduction of pubescence on the dorsum of mesosoma, reduced number of setae on subcosta of submarginal vein (to one seta only), first gastral (second metasomal) segment with raised edges, unusually short tarsi, short frontal sulcus, and weak sexual dimorphism, including antennae ( Fig. 4 View FIGURE 4 , here; Bouček 1972). Although it was described as having the dorsum of mesosoma without setae, a pair of very short setae is situated near the posterior edge of the mesoscutellum ( Fig. 4E View FIGURE 4 , inset). The propodeum characters were not mentioned among the diagnostic features of the genus; however, it was mentioned that the propodeum is “distinctly constricted to a nucha” and with a “vestigial median carina”. The actual morphology of M. diparoides suggests that the median area of propodeum is indeed bearing a median strip delimited laterally by fine carinae ( Fig. 4E View FIGURE 4 ). Another interesting character of this genus is the elongate lower face with long genae ( Fig. 4D View FIGURE 4 ) resembling to some extent the face of the Pediobius species described below. One of them, P. marjoriae , is also associated with ants, similarly to M. diparoides . The general habitus, the wide propleural flanges and the prepectus overlapped by mesosternal projection suggest the similarity of Myrmokata with Pediobius , whereas the other diagnostic characters of the genus may be secondary modifications associated with its parasitism in ants. However, the wide propleural flanges and prepectus overlapped by mesosternal projection are characteristics shared with the genera Horismenus and Microdonophagus , and the males of the latter demonstrate numerous morphological reductions associated with life in ant nests. Possible relationships between Myrmokata and Horismenus were also suggested by Bouček (1972). So, the status of Myrmokata may remain unrevised until more data (for instance, molecular) are available.













No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Pediobius Walker, 1846
| Gumovsky, Alex 2018 |
Rhynchentedon
| Girault 1919: 166 |
Pediobomyia
| Girault 1913: 155 |
Pleurotropis Förster, 1856: 78
| Forster 1856: 78 |