
Diognetus Distant, 1904
|
publication ID |
https://doi.org/10.37520/aemnp.2023.001 |
|
publication LSID |
lsid:zoobank.org:pub:3F2C90B1-6EA1-4B38-A218-C314D09F6E00 |
|
persistent identifier |
https://treatment.plazi.org/id/03F587DF-FFF1-E109-0F67-6C21FA34F996 |
|
treatment provided by |
Felipe |
|
scientific name |
Diognetus Distant, 1904 |
| status |
|
Genus Diognetus Distant, 1904 View in CoL View at ENA
Diognetus Distant, 1904b: 431–432 View in CoL (new genus). Type species by original designation: Diognetus intonsus Distant, 1904b View in CoL from Sri Lanka; SCHUH (1995): 760 (catalog); SCHUH (2002 –2013) (online catalog); CH ḖROT et al. (2017): 93–94 (diagnosis).
Diophantus Distant, 1909: 510 (new genus). Type species by original designation: Diophantus literatus Distant, 1909 View in CoL from Sri Lanka (synonymized by CARVALHO 1952: 87).
Gorna Poppius, 1914 b: 130–131 View in CoL (new genus). Type species by original designation: Gorna pilosa Poppius, 1914 b View in CoL from Java, Indonesia (synonymized by CH ḖROT et al. 2017: 94).
Diplotrichiella Poppius, 1915 a: 65–66 View in CoL (new genus). Type species by original designation: Diplotrichiella rufescens Poppius, 1915 a View in CoL from Tamil Nadu, India. New junior subjective synonym.
Diplotrichiella: SCHUH (1995) View in CoL : 761 (catalog), SCHUH (2002 –2013) (online catalog).
Yamatolygus Yasunaga, 1994: 119 View in CoL (new genus, key to Japanese species). Type species by original designation: Yamatolygus pilosus Yasunaga, 1994 View in CoL (= Diognetus yamato Yasunaga nom. nov.) [junior secondary homonym of Diognetus pilosus ( Poppius, 1914) ] from Honshu, Japan. New junior subjective synonym.
Yamatolygus: SCHUH (1995) View in CoL : 972 (catalog); KERZHNER & JOSIFOV (1999): 182 (catalog); YASUNAGA (2001): 280 (diagnosis); SCHUH (2002 –2013) (online catalog); OH et al. (2018): 482 (faunal list), 484 (generic key); AUKEMA (2018) (online catalog).
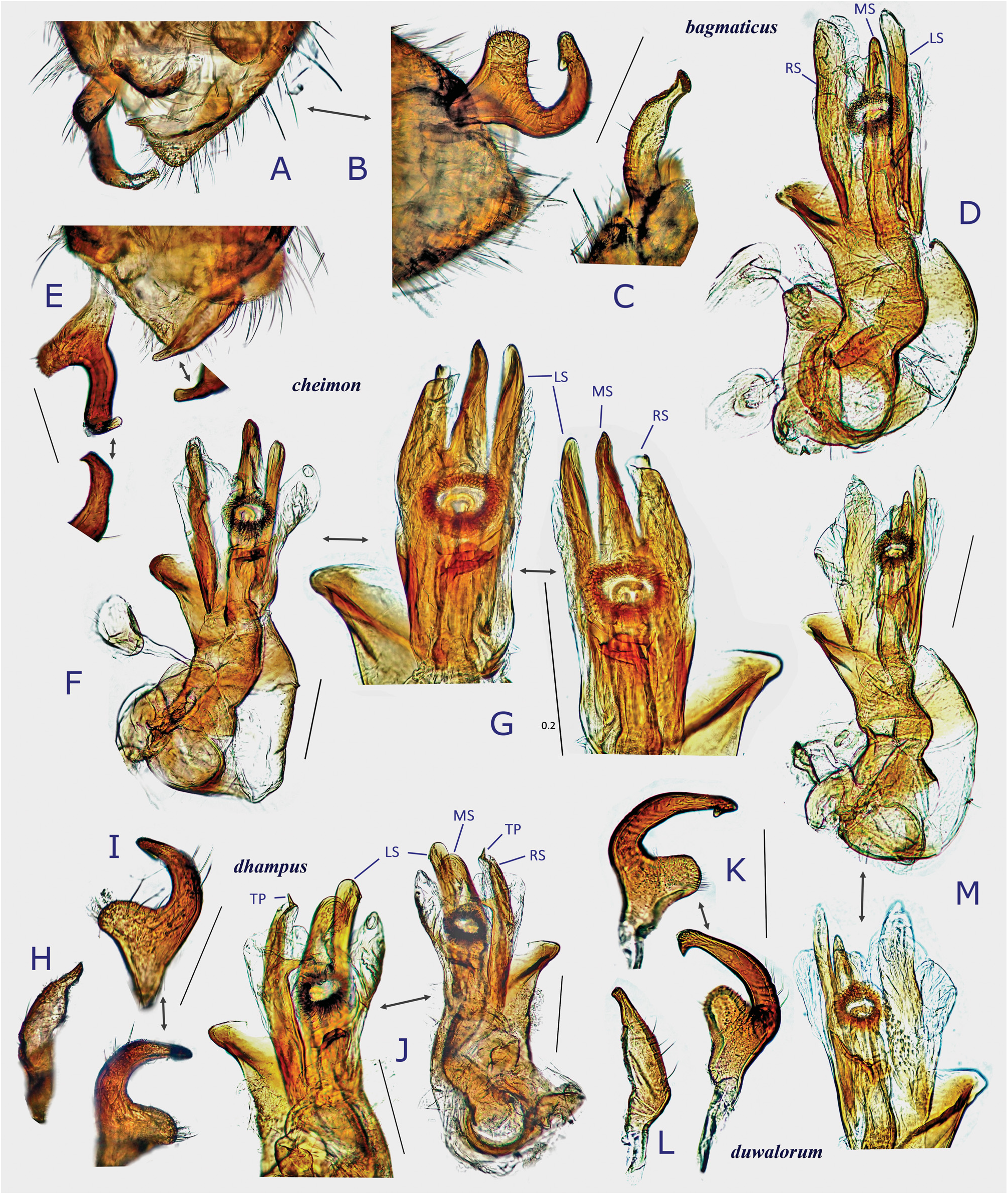
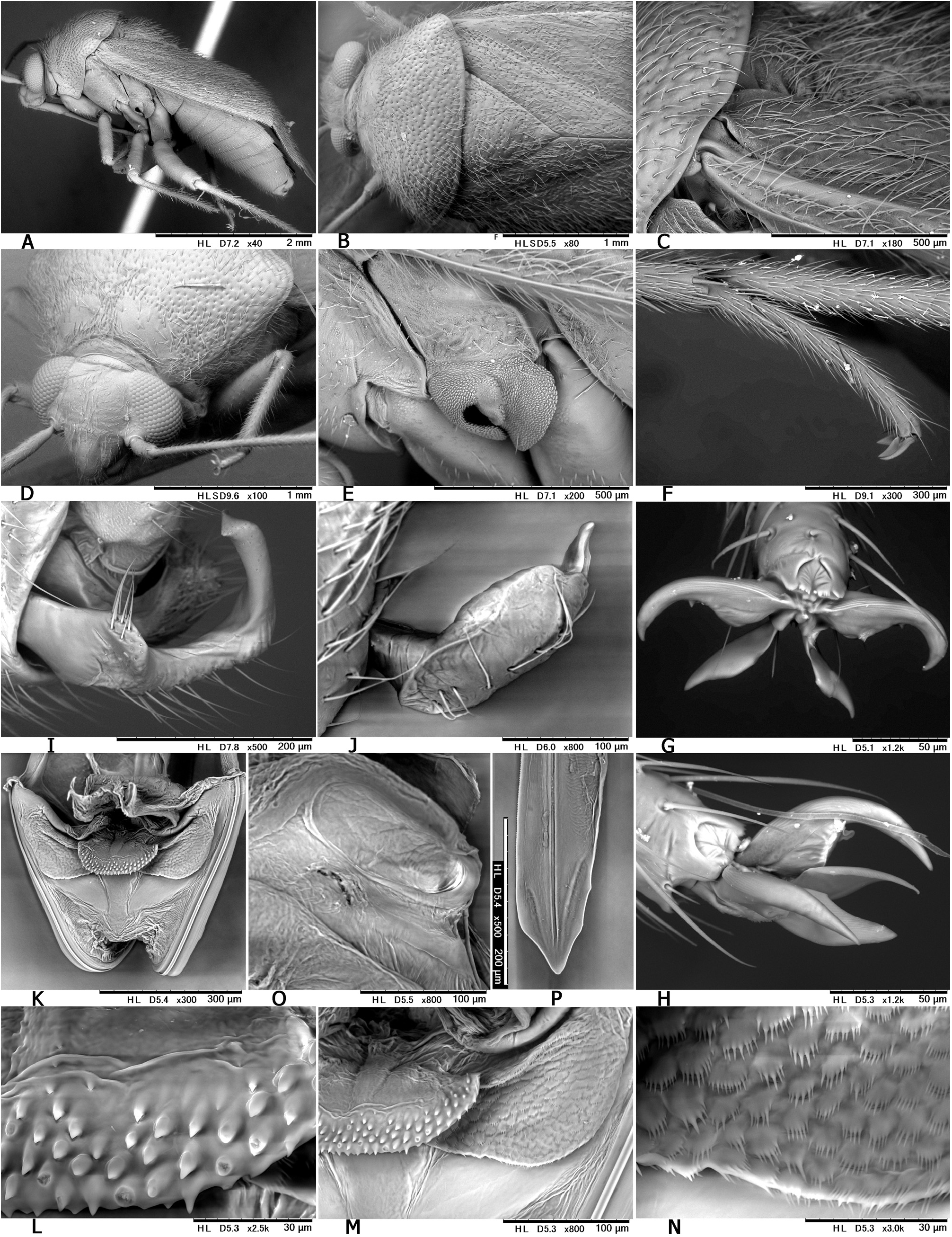
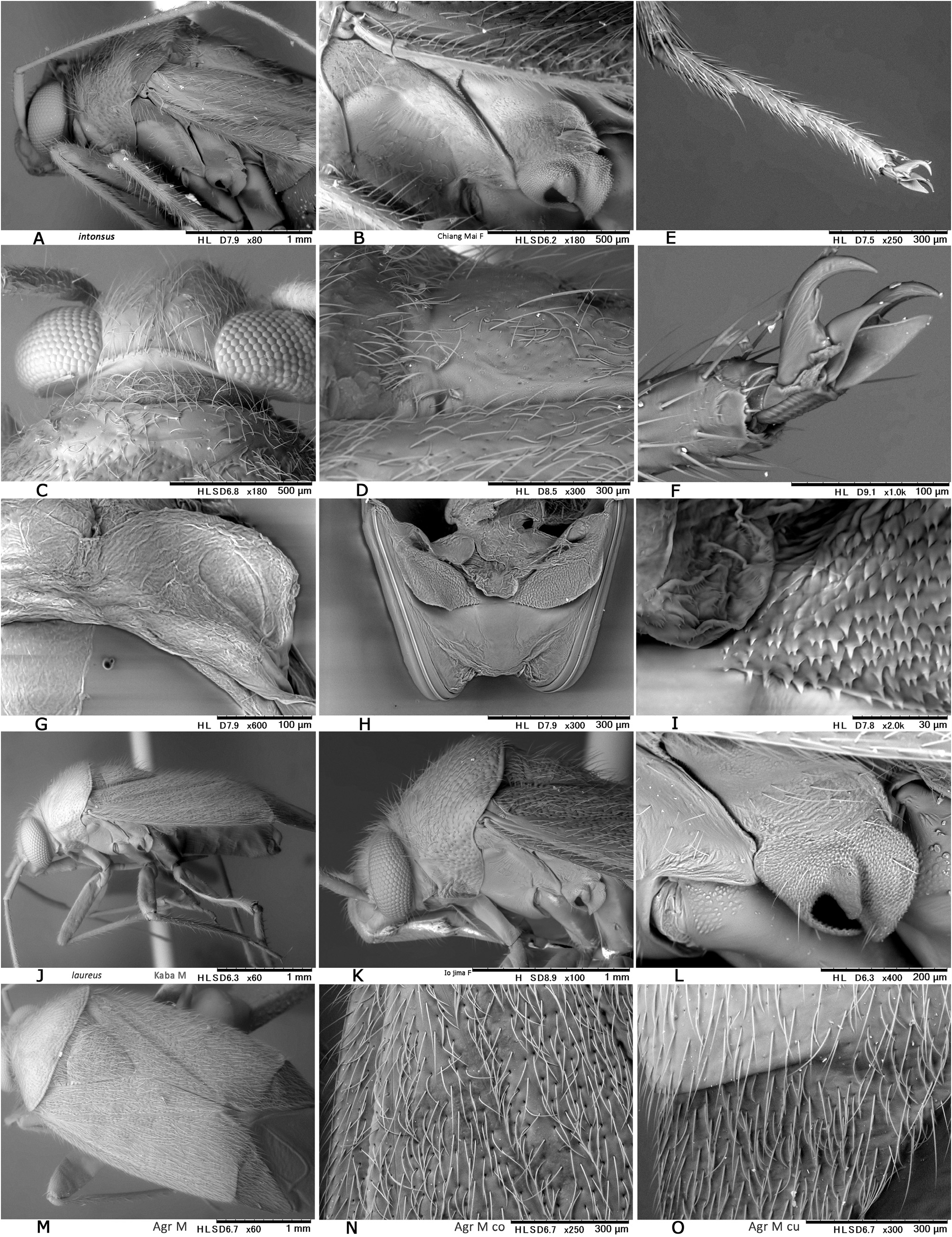
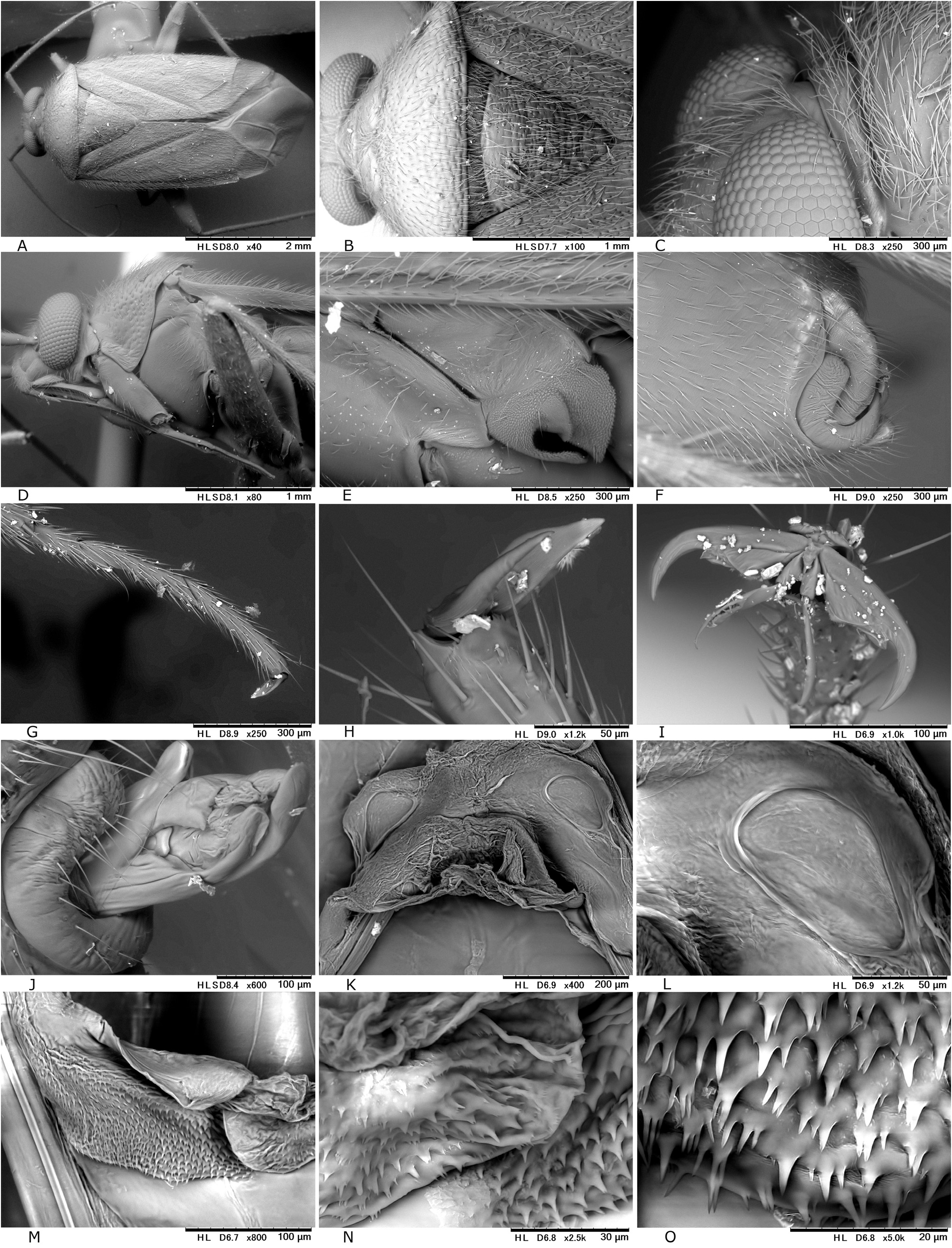
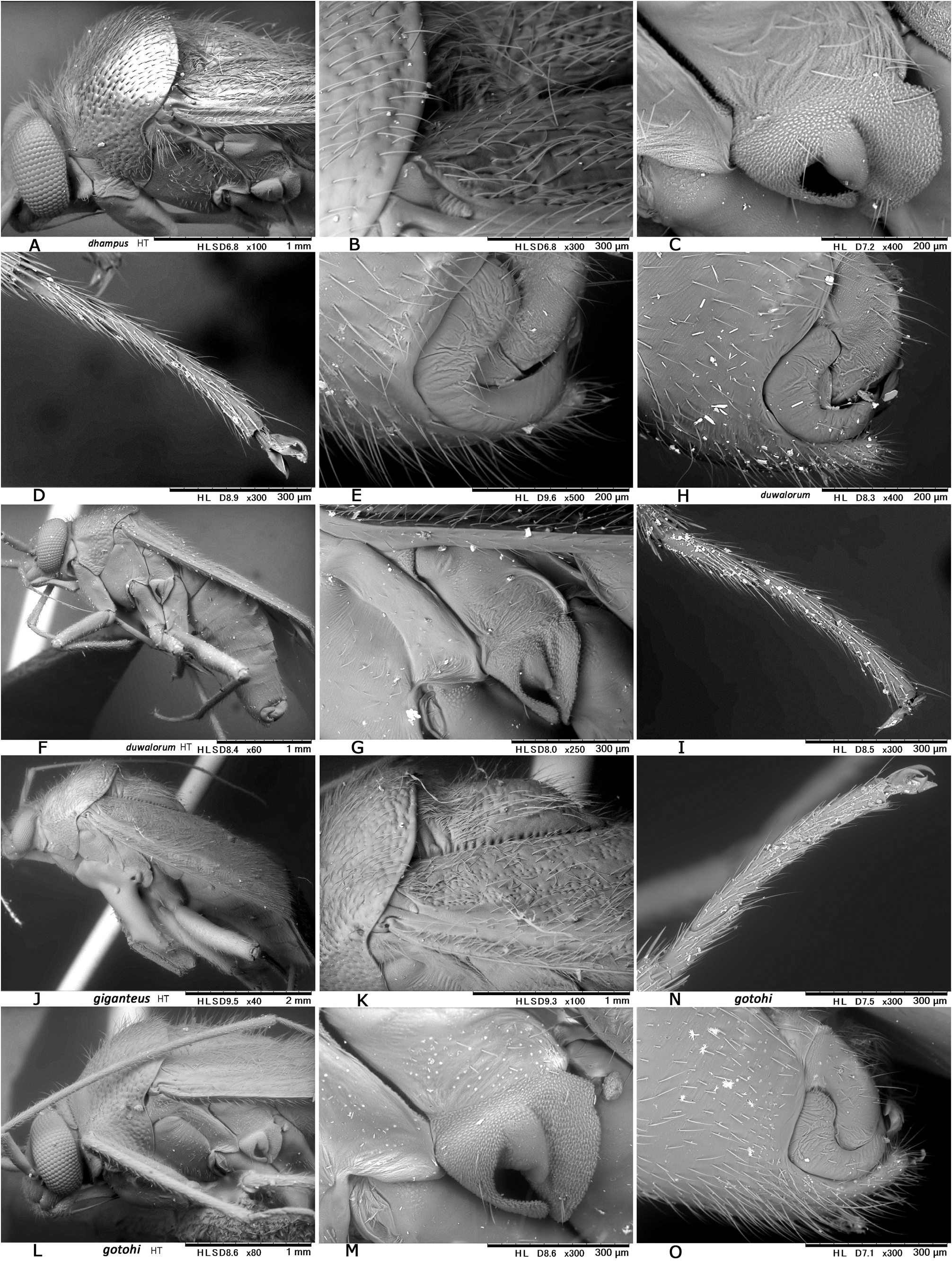
Revised diagnosis. Body ovoid to elongate-oval, rather small to moderate-sized ( 3.8–6.8 mm in total length, 1.6–3.3 in maximum width), with two or three types of vestiture (brownish semierect setae, silvery or woolly reclining setae and/or fuscous upright setae); as in many other mirines, sexual dimorphism detectable in body shape (usually more elongate in J), vertex width (narrower in J) and length of antennomeres (sometimes shorter in ♀); basic coloration brown, dark brown, and/or reddish-brown; dorsum weakly shining, uniformly punctate on pronotum, scutellum and hemelytron; head short, vertical, with brown upright setae; eyes contiguous to pronotal collar; vertex clearly carinate basally, sometimes weakly depressed or sulcate mesally; antenna generally slender and almost linear, slightly shorter than body, with segment I more or less constricted at middle; labium rather thick, variable in length (reaching mesocoxa to exceeding apex of metacoxa); pronotum rather inflated posteriorly, uniformly punctate except on calli, not carinate laterally, usually with both silvery reclining setae and brown semierect setae; pronotal collar grayish, matte, flattened dorsally, as thick as or slightly thicker than first antennal segment; punctures on propleuron sometimes reduced; scutellum weakly or moderately arched, densely pilose, sparsely or uniformly punctate, often transversely rugose; scent efferent system pale, large, about as thick as an eye in lateral view; hemelytron matte or weakly shining, distinctly punctate, with two or three types of vestiture; posterior margin of membrane vein nearly right-angled; femora and tibiae with pale brown, long, upright setae; tibial spines brown; each tarsomere I shorter than II or III; pretarsus with rhombic, developed parempodia and distinct pulvilli; male genital segment small, with pointed apex; sensory lobe of left paramere tumid, strongly produced dorsally; vesica (endosoma) small-sized, with three rod-like sclerites (namely, median lobal sclerite ‘MS’, right lobal-sclerite ‘RS’ and left lobal sclerite ‘LS’, cf. Figs 8D, F− G, J, M View Fig ) but partly reduced, elongate, spinulate or branched in some species and tiny triangular process (TP) present at apex of RS; female bursa copulatrix with small, ovoid sclerotized rings that are separated from each other; posterior wall with rather narrow, subtriangular interramal lobe and small dorsal structure; and ovipositor (gonapophysis) I rather slender, with rounded or blunt apex. Further diagnostic characters were suggested by CH ḖROT et al. (2017), POPPIUS (1915) as Diplotrichiella and YASUNAGA (1994) as Yamatolygus . The 3rd to final (5th) instar immature forms uniquely have the four transversely arranged dark or reddish spots on the dorsal anal ridge of abdominal tip (e.g., Figs 3F View Fig , 5D View Fig , 13I View Fig 13 ). Although this character has been confirmed for four species only, it may be a synapomorphy for the genus. Interestingly, the unique spots were found to remain in the newly emerging adults when the bodies are not fully chitinized ( Figs 5H, M View Fig ).
Biology. Diognetus members are basically arboreal, associated with various broadleaf trees (mostly found on the fruits, flower buds and inflorescences). The breeding host plants were confirmed (with associated immature forms) for the following congeners: Diognetus cheimon sp. nov. – Eurya emarginata (Thunb.) Makino and E. japonica Thunb. (Pentaphylacaceae) ; D. styrax sp. nov. – Styrax formosanus Matsum. ( Styracaceae ); D. laureus sp. nov. – Machilus thunbergii Sieb. et Zucc. (Lauraceae) ; D. vernus sp. nov. – Castanopsis sieboldii (Makino) Hatus (Fagaceae) , Eurya emarginata and E. japonica ; D. yamato – Symplocos coreana (H.Lév.) Ohwi and S. sawafutagi Nagam. (Symplocaceae) . Third to fifth instar immature forms of three Japanese species, D. cheimon sp. nov. ( Figs 5A–E View Fig ), D. laureus sp. nov. ( Figs 14F–I View Fig ) and D. vernus sp. nov. ( Figs 5I–K View Fig ), were reared on (commercially available) fermented milk beverage (cf. Fig. 14G View Fig ) and successfully developed into adults in laboratory tests (cf. MIYAZAKI et al. 2020).
The majority of congeners are assumed to be thermophilic and have a bivoltine or multivoltine life cycle; species inhabiting deciduous forests in temperate climate zone (e.g., D. flavigenis , D. yamato ) appear to be univoltine. The late instar immatures of two Japanese Eurya -feeding species, D. cheimon and D. vernus , were most frequently found in December, and the adults emerge until late January, possibly due to the host trees yielding fruits, or providing essential diet during the winter season.
Diognetus species are considered to be principally nocturnal, as UV lighting method has frequently yielded a number of specimens at night. In laboratory observations for D. cheimon and D. vernus , the reared adults became active and often took flight in the cages during midnight. A specimen (misidentified as D. intonsus ) was collected by fogging Neonauclea sp. (Rubiaceae) in Papua New Guinea ( CH ḖROT et al. 2017). In and around tropical or subtropical forest zones, it is difficult to sample by sweepnetting or beating method and these mirids in all likelihood inhabit canopy.
Distribution. Australia †, India, Indonesia ( Java, Sumatra †), Japan, Korea, Malaysia †, Nepal †, Papua New Guinea, Philippines †, Sri Lanka, Taiwan †, Thailand † ( YASUNAGA 2001, CH ḖROT et al. 2017, OH et al. 2008; † = confirmation by present study). Now known widely from the whole Oriental Region, Indo-Pacific across the Wallacea, and the eastern Palearctic Region; records from Australia and PNG are based on undetermined congeneric specimens (see discussion below).
Discussion. The genus Diognetus is characterized by the above diagnostic characters. Nonetheless, Poppius’ original descriptions of Gorna (in 1914) and Diplotrichiella (in 1915) also conform to Argenis Distant, 1904 (e.g., CH ḖROT 1997) ( Figs 24A–B View Fig ) and Tinginotum Kirkaldy, 1902 as well (e.g., CARVALHO 1987, EYLES 2000, YASUNAGA 1999) ( Figs 24C–G View Fig ). In addition, Tinginotopsis Poppius, 1915 (in spite of a medial hump-like protuberance on pronotum, cf. Fig. 37D View Fig ) is also similar in overall appearance to Diognetus . These nominal genera share the following characters: Dorsal main vestiture (particularly on pronotum and scutellum) sometimes very long ( Figs 24C, F View Fig ), including at least two types of setae (cf. Fig. 23 View Fig ); vertex carinate basally, weakly or faintly with longitudinal mesal sulcation (cf. Figs 24C View Fig , 28D View Fig , 30C View Fig ); eyes, particularly in male, enlarged (possibly for their nocturnal activities, as a number of specimens of Argenis and Tinginotum were collected by UV lighting at night, e.g. Figs 24A, C View Fig ), contiguous to pronotal collar; labium reaching or exceeding middle coxa; antennal segment I more or less constricted medially; pronotal calli usually flat; pronotal disc and scutellum with densely distributed, clear punctures; large areolar cell on membrane rounded apically; male genital segment parameres and vesica small-sized, comparing with body size (e.g., Figs 28A View Fig , 37I View Fig ); left paramere with developed, inflated sensory lobe and hooked apex of hypophysis; and sclerotized ring ovoid to elongate-oval, narrow-rimmed (e.g., Figs 25K− L View Fig ).
The genera mentioned above (and Tolongia Poppius, 1915 as well) can be separated from each other by the below key. Incidentally, Tinginotopsis is very close to Tinginotum and its validity is sometimes questioned (cf. EYLES 2000); the former genus can be separated from the latter only by presence of the hump-like protuberance on the pronotal disk ( Fig. 37D View Fig ). Further, some members of Tinginotum exhibit great similarity to Argenis species (e.g., T. perlatum Linnavuori, 1961 , Fig. 27G View Fig ). A comprehensive revision on Argenis-Tinginotopsis-Tinginotum complex is required on worldwide basis. As the genera include numerous species known almost throughout the Old World tropics and subtropics, however, a definitive classification for these outgroup taxa is beyond the scope of the present work.
We have recognized two lineages, or groups of species in Diognetus . Four congeners, D. flavigenis , D. minusculus , D. puspae and D. yamato , possibly constitute a monophyletic group, sharing the following characters assumed as autapomorphies: Basal part of left paramere (sensory lobe) narrowly and triangularly projected; vesical right lobal sclerite well-sclerotized, often forming a spicule; and sclerotized ring relatively thick-rimmed.All other Diognetus species generally have the basal protuberance of the left paramere widened and squared or rounded, the vesical right lobal sclerite similar in shape to two other sclerites (LS and MS), and the sclerotized ring thin-rimmed.
Incidentally, there are some additional specimens from W. Sumatra (00419677, CNC), Java (00419720, CNC) and Sulawesi, Indonesia (TYCN), Papua New Guinea (FC n° 3989, ISNB), and Luzon, Philippines (00419676, CNC); most of them are teneral or damaged, and we cannot ascertain their exact identities. It is evident that more undescribed congeners are present in tropical Asia. Therefore, a detailed phylogenetic discussion is not provided as being premature, and our species key, based mainly on the male genitalic structures, is provisional. Diognetus giganteus sp. nov. and D. pilosus (Poppius) , each known only by a single female specimen, are excluded from the key.
The following specimens from Australia (cf. Fig. 7D View Fig ) and Papua New Guinea obviously belonging to Diognetus will be treated in a separate work:
AUSTRALIA: QUEENSLAND: Mcllwraith Range,Cap York, rocky shrub, -13.88428, 143.23181, 340 m ( 1,115 ft), 29 Jun 1932, Darlington, 2 JJ ( CNC) ( AMNH _ PBI 00419632–33); North Queensland, 1.6 km S of Kuranda Turnoff on Hwy 1, -16.84047, 145.61436, 386 m ( 1,266 ft), 26 Apr 1990, J. Heraty, 1 ♀ (00419637) ( CNC). PAPUA NEW GUINEA: MILNE BAY PROVINCE: Normanby Island, Mt. Pabinama, No. 2 (Fifth Archbold Expedition to New Guinea), -10.07971, 150.99891, 820 m ( 2,690 ft), 3–5 May 1956, Fifth Archbold Exped. New Guinea, 1 J (00419628) ( CNC); Biniguni, Gwariu River, -9.66670, 149.26670, 150 m ( 492 ft), 27 Jul–14 Aug 1973, G. M. Tate, 1 J (00419629) ( CNC).








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Diognetus Distant, 1904
| Yasunaga, Tomohide, Schwartz, Michael D. & Chérot, Frédéric 2023 |
Diplotrichiella : SCHUH (1995)
| SCHUH R. T. 1995: 761 |
Yamatolygus : SCHUH (1995)
| OH M. & YASUNAGA T. & DUWAL R. K. & LEE S. 2018: 482 |
| YASUNAGA T. 2001: 280 |
| KERZHNER I. M. & JOSIFOV M. 1999: 182 |
| SCHUH R. T. 1995: 972 |
Yamatolygus
| YASUNAGA T. 1994: 119 |
Diophantus
| CARVALHO J. C. M. 1952: 87 |
| DISTANT W. L. 1909: 510 |
Diognetus
| SCHUH R. T. 1995: 760 |
| DISTANT W. L. 1904: 432 |