
Diognetus laureus, Yasunaga & Schwartz & Chérot, 2023
|
publication ID |
https://doi.org/10.37520/aemnp.2023.001 |
|
publication LSID |
lsid:zoobank.org:pub:3F2C90B1-6EA1-4B38-A218-C314D09F6E00 |
|
persistent identifier |
https://treatment.plazi.org/id/60B57FD3-00CB-442C-8E52-4873BD9506E1 |
|
taxon LSID |
lsid:zoobank.org:act:60B57FD3-00CB-442C-8E52-4873BD9506E1 |
|
treatment provided by |
Felipe |
|
scientific name |
Diognetus laureus |
| status |
sp. nov. |
Diognetus laureus sp. nov.
( Figs 1D View Fig , 10H–K View Fig , 12G− I View Fig , 13 View Fig 13 , 23 View Fig , 30J− O View Fig , 31 View Fig ) Yamatolygus sp. 1 : MIYAZAKI et al. (2020): 66, table 1 (life span, Japanese name).
Type material. HOLOTYPE: J, JAPAN: KYUSHU: Nagasaki Pref., Nagasaki City, Yotsue-machi, Agri-Hills Park, 32.809700, 129.797000, sweeping fruits of Machilus thunbergii Siebold et Zucc. (Lauraceae) , 25 Jun 2019, T. Yasunaga ( AMNH) ( AMNH _ PBI 00380751). PARATYPES: JAPAN: KYUSHU: Fukuoka Pref., Yame City, Mt. Shakadake, 33.20, 130.84, UV lighting, 6 Jul 2019, 1 ♀, T. Nozaki ( WCF); Nagasaki Pref., Nagasaki City,Tateyama, Mt.Konpira, sweeping flower buds of Eurya japonica , 32.764888, 129.879500, 24 Dec 2021, T.Yasunaga, 1♀ ( TYCN); Nagasaki City, Nomozaki, Kabashima Island, 32.556555, 129.777977, sweeping fruits of Machilus thunbergii , 1 Jun 2019, T. Yasunaga, 1 J [4th instar immature form reared then emerging on Jun 5, 2019, and mounted on Jun 16] ( TYCN); same data, except for date, 6 Jun 2021, T. Yasunaga, 1 J 3 ♀♀ ( TYCN); same data, from 32.556161, 129.778000, inflorescence of Cinnamomum japonicum Sieb. , 6 Jun 2021, T.Yasunaga, 1 ♀ [5th instar immature form reared then emerging on same day and dead on Jun 12] ( TYCN); Nagasaki Pref., same data as for holotype, 2 JJ 2 ♀♀ ( TYCN); Nagasaki City,Iojima Island (lighthouse-park), 32.714177, 129.761155, sweeping flowers and fruits of Eurya emarginata , 27 Dec 2019, T. Yasunaga & K. Tanaka, 1 ♀ [5th instar immature form reared then emerging on Jan 1, 2020] ( TYCN); Nagasaki City, Tateyama; Mt. Konpira, 32.765247, 129.877073, sweeping flowers and fruits of Machilus thunbergii , 19 Jun 2021, T. Yasunaga, 6 JJ ( CNC, TYCN); same data, except for detailed collecting site, 32.7640, 129.8787, 1 ♀ ( TYCN); Nagasaki Pref., Omura City, Kushima, Ryujin Island (Shrine), 32.895166, 129.952900, on flower of evergreen broadleaf, 4 Jul 2022, T.Yasunaga, 1 J ( TYCN). SHIKOKU: Kochi Pref., Otoyo Town, Osugi, 33.76, 133.66, on Ternstroemia gymnanthera (Wight &Arn.) Bedd. (Pentaphylacaceae) , 14 Jul 2004, M. Takai, 1 ♀ ( TYCN); Tosa-Shimizu City, Cape Ashizuri, 32.73, 133.00, 22 May 1999, T. Befu, 1 J ( TYCN).
Additional material examined (teneral specimens): JAPAN: KYUSHU: Kumamoto Pref., Amakusa City, Uonuki Town, 32.256, 129.999, UV lighting, 12 Jun 2015, T. Nozaki, 1 ♀ ( WCF); Nagasaki Pref., Nagasaki City, Kabashima Island, from 32.556161, 129.778000, inflorescence of Cinnamomum japonicum Sieb. , 6 Jun 2021, T. Yasunaga, 1 ♀ [5th instar immature form reared then emerging on Jun 9 and dead on Jun
12] ( TYCN); same locality and plant as for holotype, 6 Jun 2021, T. Yasunaga, 2 JJ [5th instar immature form reared then emerging on Jun 10–11 and dead on Jun 15, Jun 20] ( TYCN).
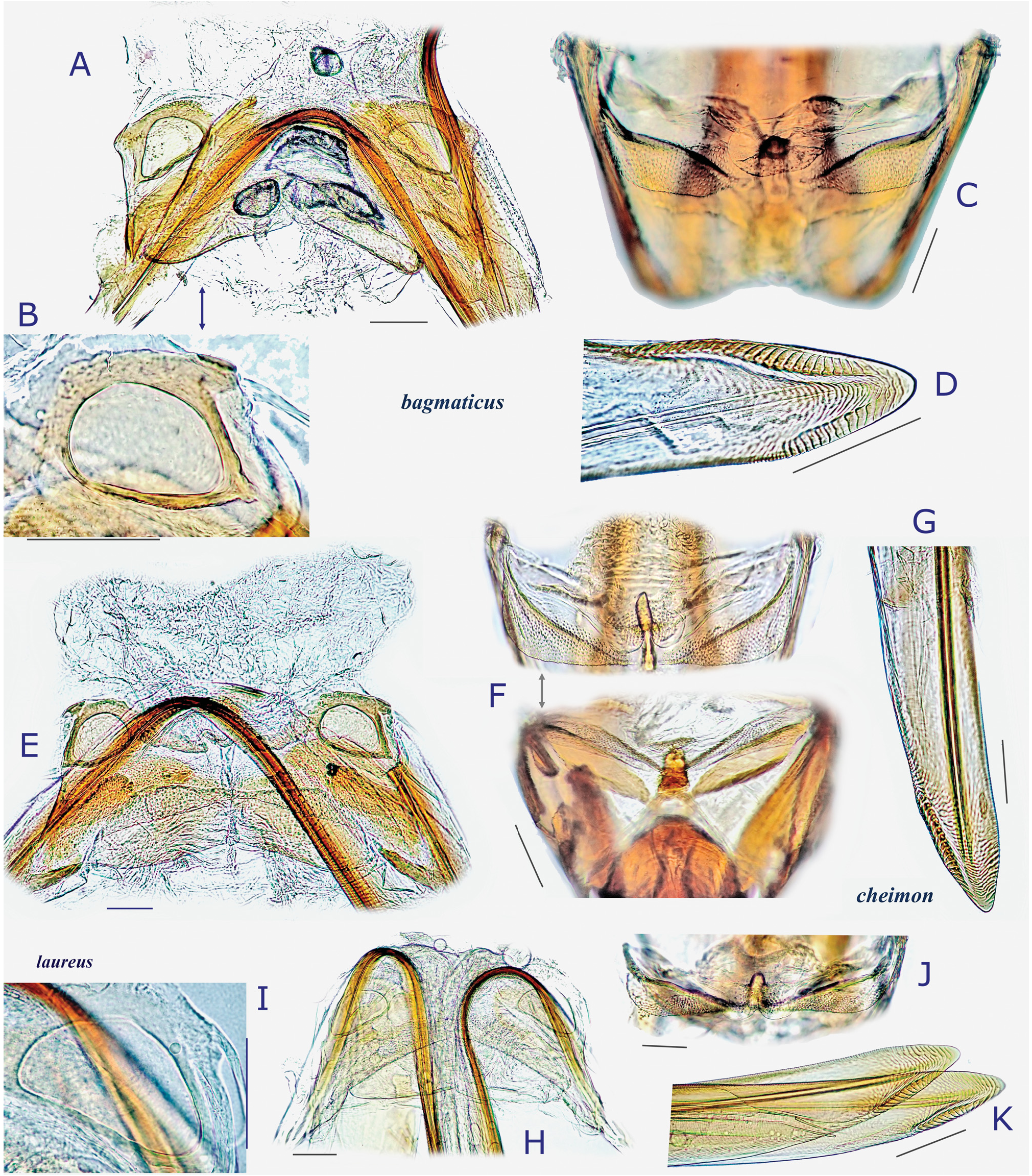
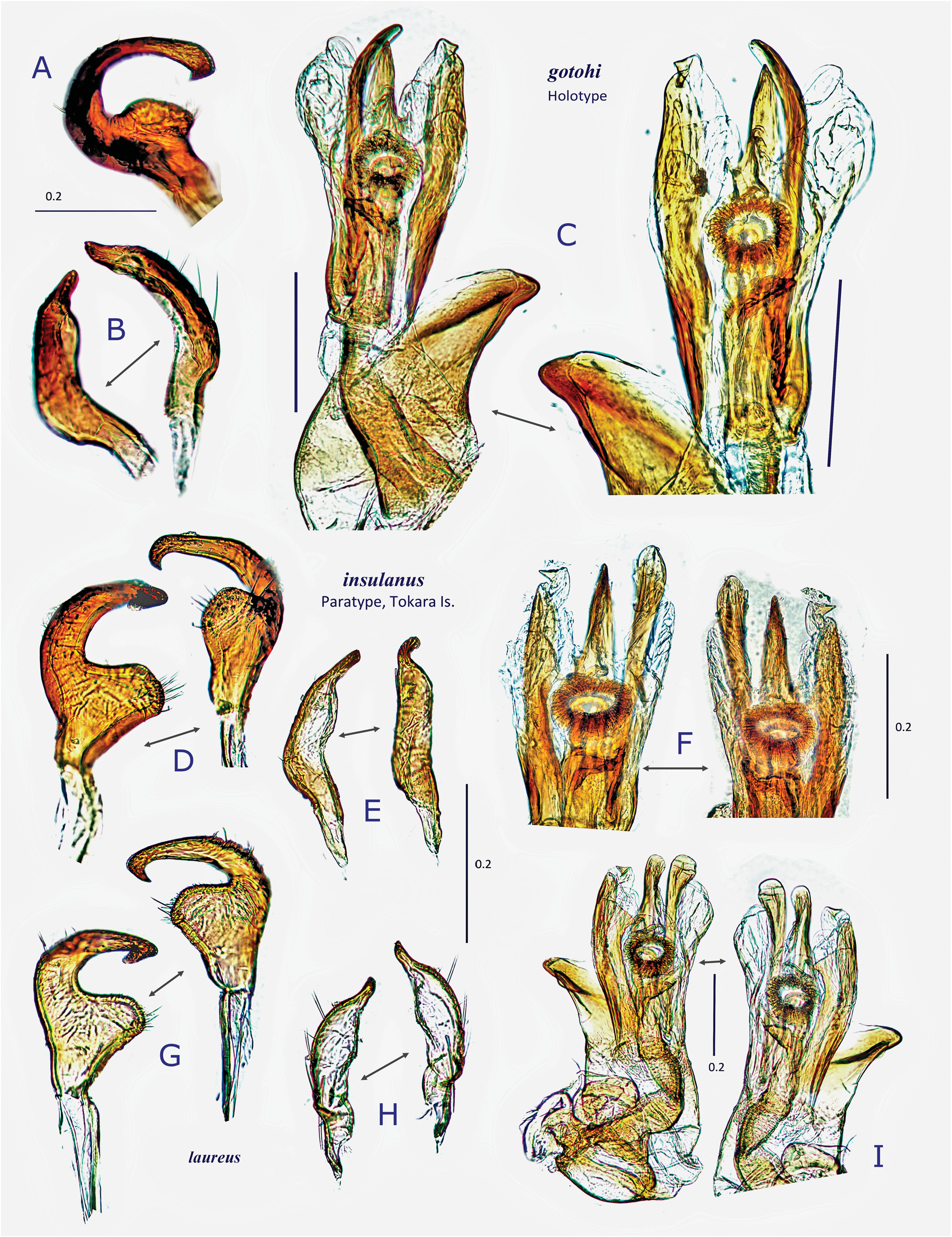
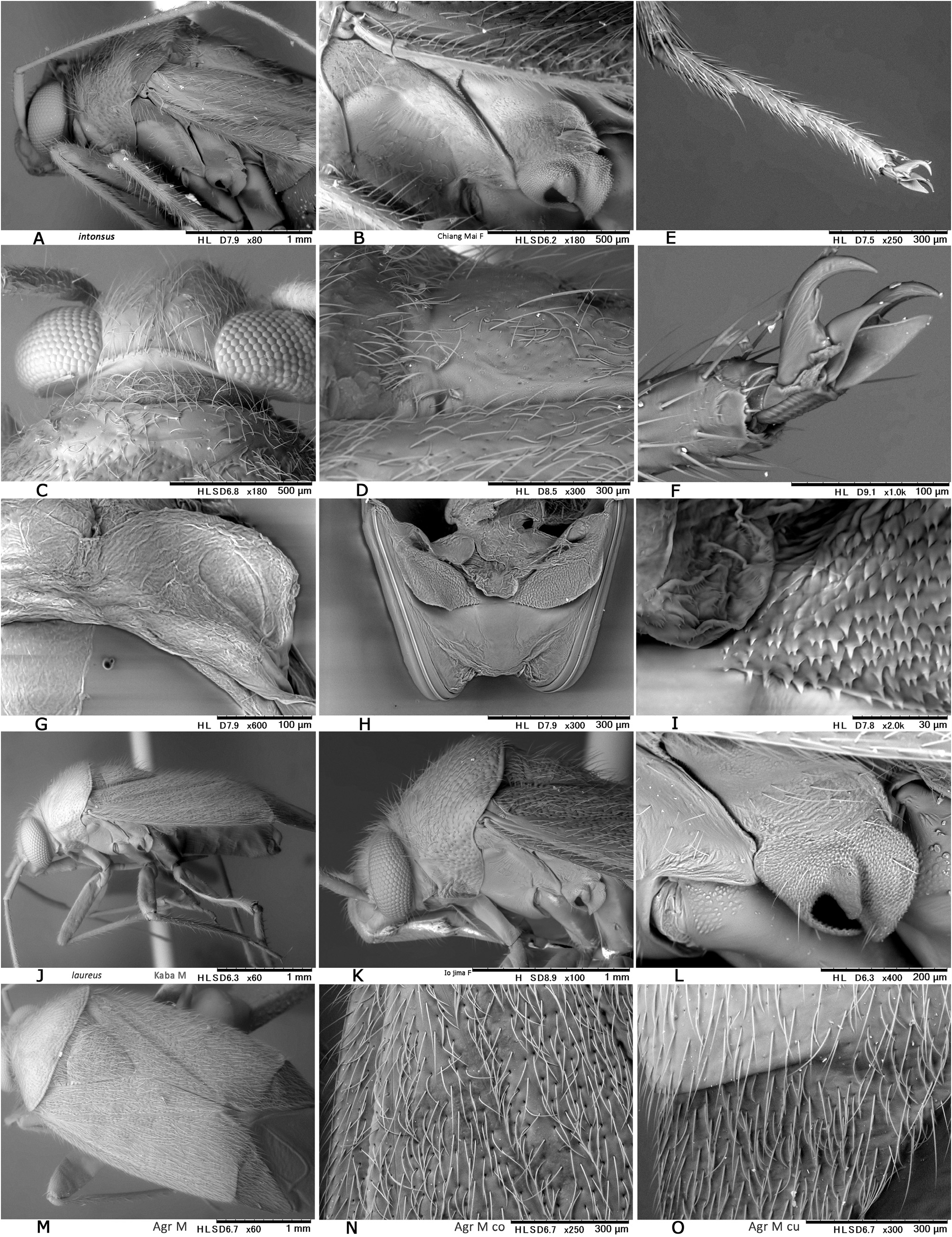
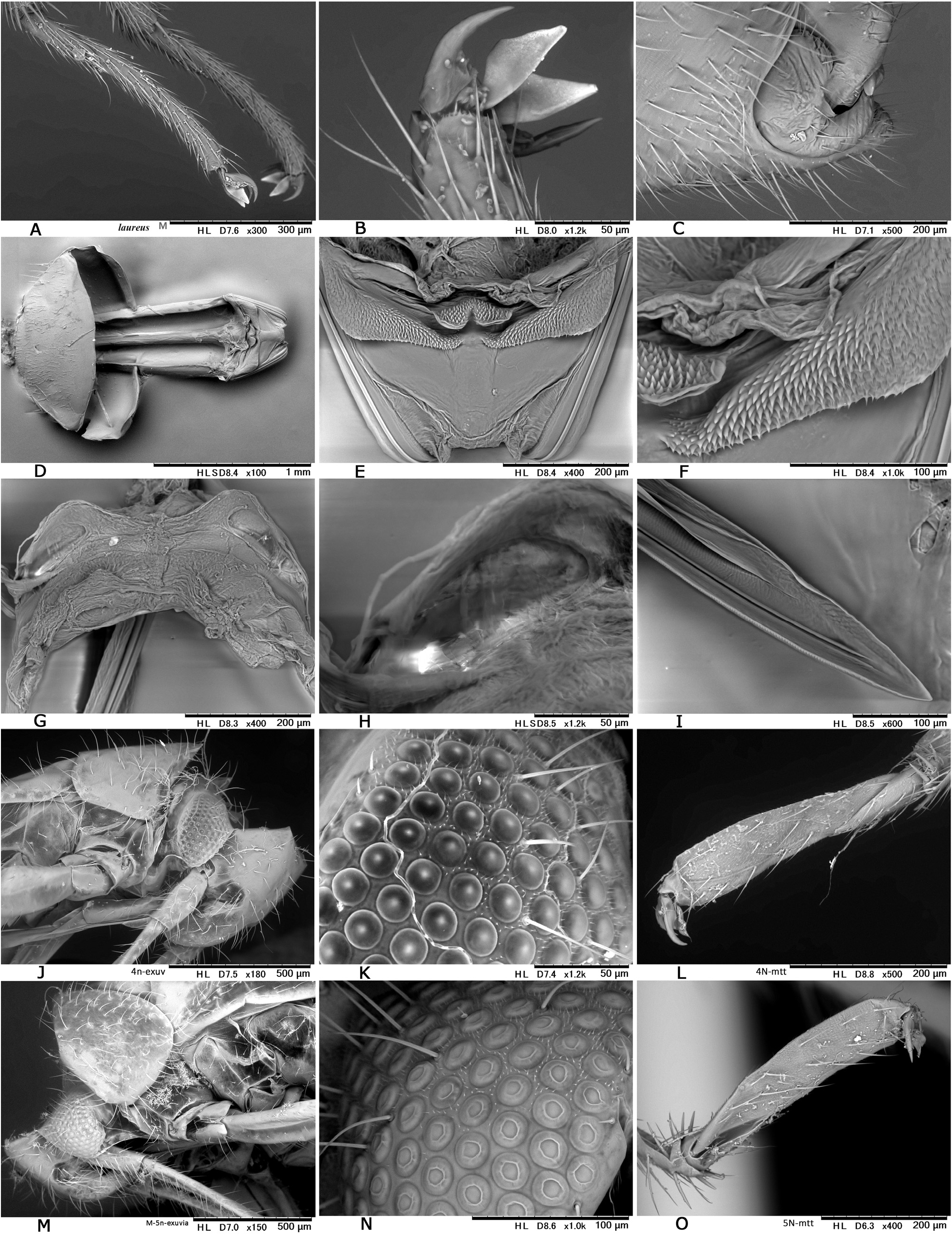
Description. Body elongate ovoid, relatively small, 4.2–5.1 mm in total length. COLORATION: Body including ventral surface generally pale reddish-brown ( Figs 13A− C, E View Fig 13 ) (fresh specimens often bright red, cf. Fig. 13D View Fig 13 ); dorsum without mottled pattern. Antennae pale reddish-brown; segment I red or reddish-brown; apex of segment II uniformly pale, partly tinged with red; segment III (except for pale extreme base) and segment IV brown. Labium pale reddish-brown; apex of segment IV infuscate ( Fig. 13E View Fig 13 ). Pronotum usually brown posteriorly, with yellowish posterior margin; calli often with a pair of dark, small spots; pleura broadly reddish-brown; scent efferent system creamy yellow. Hemelytron almost uniformly pale reddish-brown, without dark maculae; membrane pale smoky brown, with pale veins and several semitransparent spots ( Figs 13A− B View Fig 13 ). Coxae and legs reddish-brown; mesofemur with two faint pale rings subapically ( Fig. 13E View Fig 13 ); tarsi brown; apical part of each tarsomere III brown. Ventral surface of abdomen reddish-brown, with paler median part. SURFACE AND VESTITURE: As in generic diagnosis; dorsal surface weakly shining, with densely distributed, brown, simple, semierect setae and silvery, reclining setae ( Figs 23 View Fig , 30N View Fig ); scutellum transversely rugose, sparsely and finely punctate; hemelytron rather matte; punctures on corium partly very sparse or missing. STRUCTURE: Vertex with a basal transverse carina, weakly arched. Antennal segment II obviously shorter than labium. Labium long, exceeding apex of metacoxa and reaching abdominal sterna IV− VI ( Fig. 30J View Fig ), in J longer than maximum width across hemelytron. Scutellum rather flat. Metathoracic scent efferent system as in Fig. 30L View Fig . Metatarsomere II slightly longer than III; pretarsal structure as in Fig. 31B View Fig ; parempodia rather long and broad. MALE GENITALIA ( Figs 12G− I View Fig , 31C View Fig ): Pygophore generally small in size ( Fig. 31C View Fig ). Left paramere with relatively narrow protuberance of sensory lobe and somewhat tapered hypophysis that is hooked at apex ( Fig. 12G View Fig ); right paramere slender ( Fig. 12H View Fig ). Vesica small-sized, with MS and LS that are similar in shape and size; RS minutely and sparsely spinulate, with small TP ( Fig. 12I View Fig ). FEMALE GENITALIA ( Figs 10H− K View Fig , 31D− I View Fig ): Sclerotized ring thin-rimmed ( Figs 10H− I View Fig , 31G− H View Fig ); posterior wall ( Figs 10J View Fig , 31E− F View Fig ) with rather small dorsal structure; interramal lobe with rounded lateral margin ( Fig. 31E View Fig ); spines on dorsal structure smaller than interramal lobe ( Fig. 31F View Fig ).
Measurements. See Table 1.
Differential diagnosis. Recognized by its generally pale reddish-brown, relatively small-sized, elongate oval body; long labium that is surpassing apex of metacoxa and longer than maximum width in J; generally small male genitalia; and thin-rimmed sclerotized ring. Most closely related to D. insulanus , from which D. laureus sp. nov. can be distinguished by the generally smaller size, pale reddish-brown basic coloration, longer labium, narrower apical part of the protuberance of the left paramere, thinner-rimmed sclerotized ring, rather rounded lateral margin of the interramal lobe, and shorter spines on the dorsal structure.
The immature forms (3rd to 5th instars) are recognized by the following features: Body generally bright red, rounded or ovoid (sometimes partly tinged with olive as in Fig. 13G View Fig 13 ); antenna long, longer than body in 4th−5th ( Figs 13F, I View Fig 13 ); and dark spot (scent gland opening) on abdominal tergum III reddish, small. By these characters, the late immature forms of D. laureus sp. nov. can be distinguished from those of D. cheimon sp. nov. and D. vernus sp. nov. (cf. Fig. 5 View Fig ).
Etymology. Named for the plant association of this new species with Lauraceae hosts; an adjective.
Biology. This new species is associated with Lauraceae evergreen broadleaf trees (flowers and fruits), Machilus thunbergii Siebold & Zucc. ( Fig. 1D View Fig ) and Cinnamomum japonicum Sieb. , on which the immature forms were found in June. The reddish general coloration both in the adults and nymphs appears to be cryptic, harmonious with the red peduncles (cf. Figs 13C, I View Fig 13 ). On the other hand, winter- -emerging population (possibly 2nd annual generation, cf. Table 2) was discovered from the flowers or fruits of Eurya emarginata (and 1 ♀ from E. japonica ) and often co-occurred with Diognetus cheimon sp. nov. and D. vernus sp. nov. These three mirids are closely related to each other and may be derived from a common ancestor that was associated with Eurya evergreen trees, although the spring− summer generation of each species utilizes different hosts.
Distribution. Japan (Shikoku, Kyushu).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.