
Zischkaia arenisca Nakahara, Willmott & Hall, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.551 |
|
publication LSID |
lsid:zoobank.org:pub:C3C851C3-0F12-412C-A15B-56F0F263CD00 |
|
DOI |
https://doi.org/10.5281/zenodo.3477356 |
|
persistent identifier |
https://treatment.plazi.org/id/83701B3C-3A99-4BFC-9E8E-18F03C47CE0B |
|
taxon LSID |
lsid:zoobank.org:act:83701B3C-3A99-4BFC-9E8E-18F03C47CE0B |
|
treatment provided by |
Plazi |
|
scientific name |
Zischkaia arenisca Nakahara, Willmott & Hall |
| status |
sp. nov. |
Zischkaia arenisca Nakahara, Willmott & Hall , sp. nov.
urn:lsid:zoobank.org:act:83701B3C-3A99-4BFC-9E8E-18F03C47CE0B
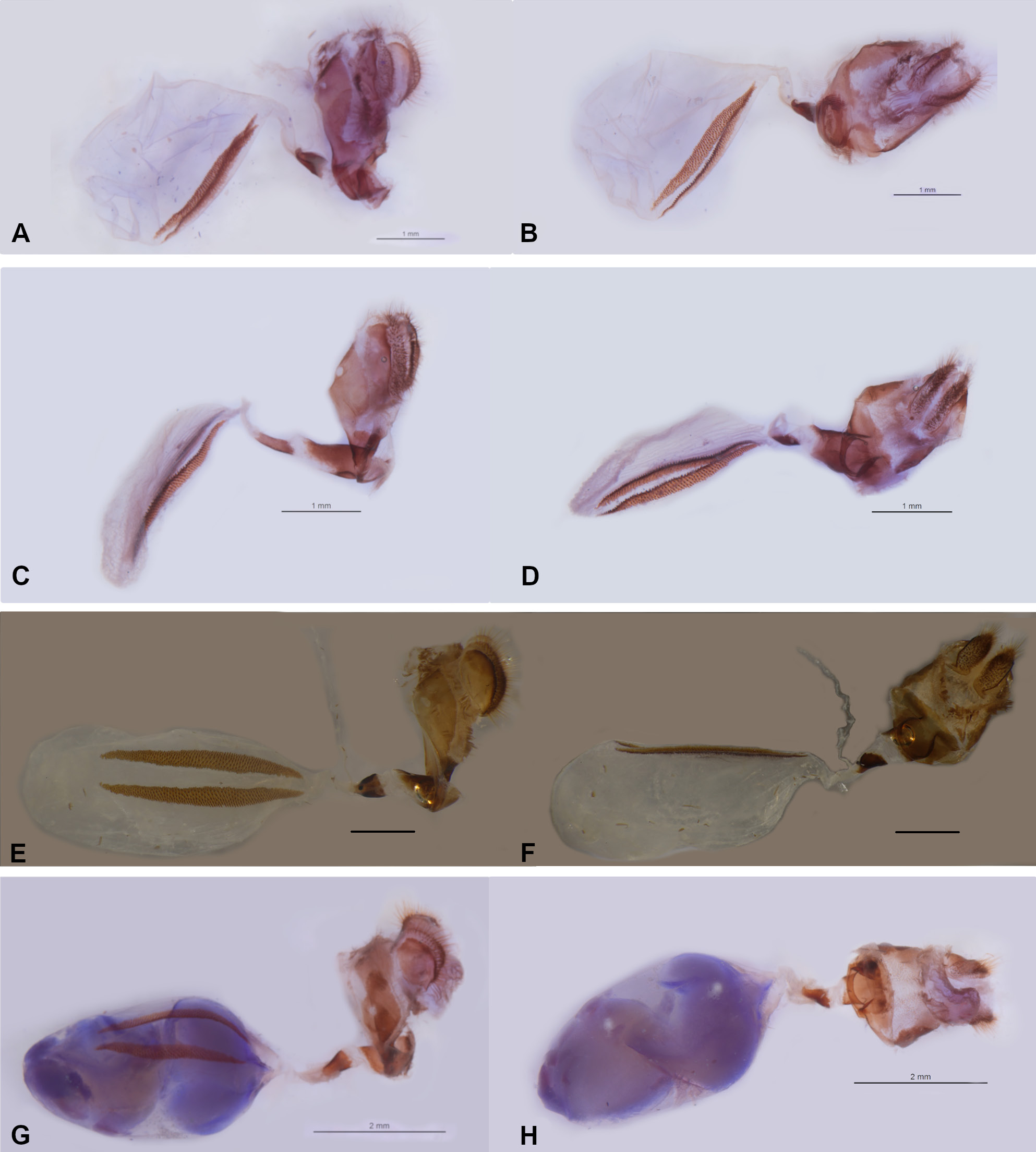
Figs 1 View Fig , 2 View Fig M–P, 4 View Fig E, 6 View Fig E–F, 9 View Fig , 14 View Fig – 15 View Fig A–B
Zischkaia sp. n. – Lamas 1997: 217; 2004: 223.
Diagnosis
Zischkaia arenisca sp. nov. is readily distinguished from all species of Zischkaia , except for Z. argyrosflecha sp. nov., by its elongate, oval VHW ocelli, whereas those ocelli are more circular and smaller in Z. pacarus , Z. amalda , Z. baku sp. nov. and Z. chullachaki sp. nov. Species in the “ saundersii clade” also possess somewhat elongate VHW ocelli, but the genitalia differences summarized above (see diagnosis for Zischkaia ), in addition to the lack of dorsal androconial scales in males, easily distinguish this species from species in the “ saundersii clade”. The male DFW and DHW androconial scales in Z. arenisca sp. nov. appear somewhat reduced and less prominent compared to those of Z. baku sp. nov. and Z. chullachaki sp. nov. Furthermore, the forewing apex is somewhat rounded in Z. arenisca sp. nov., whereas it is more angled in Z. pacarus , Z. amalda and Z. baku sp. nov. In the female genitalia, the sclerotized tube of the ductus bursae in lateral view appears to be longer in Z. arenisca than in Z. chullachaki sp. nov. and Z. pacarus .
Zischkaia arenisca sp. nov. is likely most closely related to Z. argyrosflecha sp. nov., given their similar elongate VHW ocelli, but Z. arenisca sp. nov. can be distinguished from Z. argyrosflecha sp. nov. by the following characters: the lack of silvery-purple scales on the basal side of the VHW ocelli (distad of the submedian line); the lack of or very small VHW ocellus in Rs-M 1, whereas this ocellus is well developed in VHW Rs-M 1 in Z. argyrosflecha sp. nov.; only one winglet being present on the aedeagus, whereas Z. argyrosflecha sp. nov. possesses two winglets; the shape of the aedeagus winglet is that of an obtuseangled triangle in Z. arenisca sp. nov., whereas it is an acute-angled triangle in Z. argyrosflecha sp. nov.; the saccus is longer than the ventral margin of the valva in Z. arenisca sp. nov., whereas it is shorter than ventral margin of the valva in Z. argyrosflecha sp. nov.; the ventral margin of the apical process of the valva is not concave in Z. arenisca sp. nov., whereas it is concave in Z. argyrosflecha sp. nov. The female of Z. argyrosflecha sp. nov. is unknown and thus it is not possible to compare that sex.
Etymology
This specific epithet is derived from the Spanish word ‘ arenisca ’, meaning ‘sandstone’, and is treated as a feminine noun in apposition. The name alludes to the apparent strong preference of this species to forest growing on sandstone substrates.
Type material examined
Holotype
ECUADOR • ♂; “// ECUADOR: Zamora-Chinchipe Río Nangaritza, Shaime , 4°19’S, 78°40’W 900m, xi.1997 Jiggins, C.D.// DNA voucher LEP-67941// Genitalic vial SN-17-74 S. Nakahara // Photographed by K. R. Willmott 2004// UF FLMNH MGCL-1036291//”; FLMNH, to be deposited in INABIO. GoogleMaps
Paratypes (20 ♂♂, 4 ♀♀)
ECUADOR – Zamora-Chinchipe • 19 ♂♂; 5 km S of Zurmi, Sumak Yaku mine rd.; [4°8′38″ S, 78°38′57″ W]; 1000 m; 26, 28–29 Jul. 2018; J.P.W. Hall, K.R. Willmott, J.C.R. and J.I.R. Willmott leg.; FLMNH-MGCL-297322 to FLMNH-MGCL-297340; FLMNH GoogleMaps • 1 ♀; same collection data as for preceding; FLMNH-MGCL-297321; FLMNH GoogleMaps • 2 ♀♀; km 10 Los Encuentros-El Panguí, ridge E San Roque ; [3°42′11″ S, 78°35′36″ W]; 1050 m; 18–19, 23 Jul. 2018; J.P.W. Hall, K.R. Willmott, J.C.R. and J.I.R. Willmott leg.; FLMNH-MGCL-297319 to FLMNH-MGCL-297320; FLMNH GoogleMaps • 1 ♀; same collection data as for preceding but 4 Aug. 2009; K.R. Willmott and J.P.W. Hall leg.; FLMNH-MGCL- 149623; FLMNH GoogleMaps • 2 ♂♂; Río Nangaritza, Shaime ; [4°20′ S, 78°40′ W]; 1000 m; Sep. 1997; C.D. Jiggins leg.; FLMNH-MGCL-1036290 to FLMNH-MGCL-1036291; FLMNH GoogleMaps .
Description
Male
FOREWING LENGTH. 26–27 mm (n = 2).
HEAD. Eyes naked, with grayish scales at base; frons brownish; post-genal area with lightly colored long hair-like scales and grayish scales; labial palpi with first segment covered by white and brownish long hair-like scales, in addition to some whitish scales at base; second segment almost twice as long as eye depth and covered with brown scales and hair-like scales laterally, with partially white scales and hairlike scales, dorsally with brownish long hair-like scales, ventrally with black and white hair-like scales, 3–4 times as long as segment width; third segment about one-third of second segment in length, covered with brownish scales, with small patch of creamy-white scales laterally; antennae approximately twofifths forewing length, with ca 44 segments (n = 1), distal 16–17 segments composing club, club not prominent.
THORAX. Covered with light brownish hair-like scales.
LEGS. Foreleg brownish, foretarsus slightly shorter than tibia, femur similar to tarsus in length; midleg and hindleg with femur creamy white ventrally, tibia and tarsus grayish dorsally, ocher ventrally, tarsus and tibia spined ventrally, and a pair of tibial spurs present at distal end of tibia.
ABDOMEN. Eighth sternite divided into two sclerotized patches: a thin band at base of eighth abdominal segment, and a distal broader patch.
WING VENATION. Basal half of forewing Subcosta swollen; base of Cubitus swollen; forewing recurrent vein absent; origin of M 2 nearer M 1 than M 3; hindwing humeral vein well developed.
WING SHAPE. Forewing subtriangular, apex rounded, costal margin convex, outer margin slightly convex (almost straight), inner margin straight, but rounded towards thorax near base; hindwing slightly elongate, rounded, costal margin almost straight, angled towards thorax near base, outer margin slightly undulating, inner margin slightly concave near tornus, anal lobe convex, slightly round.
DORSAL FOREWING. Ground color light brownish, distally slightly paler; black androconial scales present in middle of DFW from base to submedian area, except for immediately adjacent area around swollen Subcosta and Cubitus, in addition to area anterior to Radial about distal one-fifth of DFW; trace of submarginal band visible.
DORSAL HINDWING. Ground color similar to forewing, black androconial scales present around distal half of discal cell, extending to adjacent area, trace of submarginal band visible.
VENTRAL FOREWING. Ground color chestnut brown; submedian line invisible; dark brown narrow median line, somewhat indistinct, extending from near costa to Cu 2, slightly curved inwards, concolorous slightly sinuate submarginal line extending from apex towards tornus, but terminating half way between Cu 2 and 2A; concolorous marginal line, narrower than submarginal line, extending from apex towards tornus, but terminating half way between Cu 2 and 2A; fringe dark brownish.
VENTRAL HINDWING. Ground color similar to forewing; regular dark-brown submedian line almost straight, extending from costal to inner margin; median band almost parallel to submedian band, concolorous, similar in width, passing origin of M 3 and posterior end bent inwards in 2A-3A; submarginal band extending from apex towards tornus, anterior end occasionally fused with submedian line near costa, undulating, posterior end slightly broadening and occasionally fused to submedian line in 2A-3A; marginal line, concolorous, slightly undulating along outer margin, thinner than submarginal line; submarginal ocelli from M 1 to 2A, roughly ‘egg’-shaped with pointed end basal, black central spot ringed with orangish ring then with thin dark brownish indistinct ring, pupil consisting of scattered silver scales displaced distally from center, ocelli from M 2 -M 3, M 3 -Cu 1 and Cu 1 -Cu 2 similar in size, somewhat elongate, those in M 1 -M 2 and Cu 2 -2A about half size of these three ocelli, ocellus in Rs-M 1 a tiny orangish spot; fringe dark brownish.
GENITALIA ( Fig. 4E View Fig ). Tegumen rounded in lateral view, elongated posterior projection of tegumen developed, slightly shorter than uncus, basal half somewhat inflated, tapering posteriorly and hooked at terminal point, posterior end of ventral margin as small projection; combination of ventral arms of tegumen and dorsal arms of saccus sinuous, bent at right angle below appendices angulares, broadens towards saccus; appendices angulares present, but somewhat reduced; saccus straight, similar to uncus in length; uncus long and narrow, with sparse hair-like setae, curved ventrally, rounded at terminal point; either side of base of uncus with short hair-like setae; brachia similar to uncus in length and width, curved in lateral view, tapering posteriorly and crossing over each other near terminal point; fultura inferior (i.e., juxta) present; valva subtriangular in lateral view, apical process triangular, ventral and dorsal margin both almost straight, scarcely covered by hairy-like setae, costa developed and triangular, dorsal margin slightly sinuous; phallobase slightly shorter than half of phallus in length, curved upwards; ductus ejaculatorius visible; aedeagus straight with manica covering approximately half, winglet present on dorsal surface, distal opening located ventrally where vesica is visible.
Female
Similar to male except as follows: FW length 27 mm (n = 1): Foreleg whitish, foretarsus divided into five distinct subsegments, fifth subsegment elongate; forewing slightly more rounded, outer margin slightly convex, ground color of both wing surfaces paler.
FEMALE ABDOMEN AND GENITALIA ( Fig. 6 View Fig E–F): inter-segmental membrane between seventh and eighth tergite not pleated, but folded posteriorly of ostium bursae with its sclerotized region forming a ‘scooplike’ structure below lamella antevaginalis; lamella antevaginalis sclerotized; posterior approximate three-fourths of ductus bursae sclerotized, remainder membranous; posterior end as a circular hole in posterior view, with edge darker (presumably reflecting degree of sclerotization), surrounding sclerotized area somewhat semi-circular in posterior view; origin of ductus seminalis close to sclerotized portion of ductus bursae; eighth tergite with lateral side sclerotized and fused to lamella antevaginalis; corpus bursae roughly oval in dorsal view, extending across entire abdomen, with two signa located in middle, extending across almost entire corpus bursae, parallel to each other.
Variation
Most individuals lack an ocellus in VHW Rs-M 1, or if it is present, it is very small (much smaller than that in 2A-Cu 2). However, in one male (FLMNH-MGCL-297332) and one female (FLMNH-MGCL-297320), both barcoded (MUSM-LEP-64859 and MUSM-LEP-64857, respectively), it is rather more developed, approximately half the size of that in cell 2A-Cu 2. The size of the VHW ocelli is also slightly variable, with smaller ocelli being more rounded. The ventral ground color is darker in some specimens, in which the paler brown rings surrounding the ocelli are also more conspicuously paler. The combination of the ventral arms of the tegumen and dorsal arms of the saccus is sinuous and bent at a right angle below the appendices angulares in the male paratype, whereas it is rather straight in the holotype; the curvature of the phallobase is variable between the two dissected males.
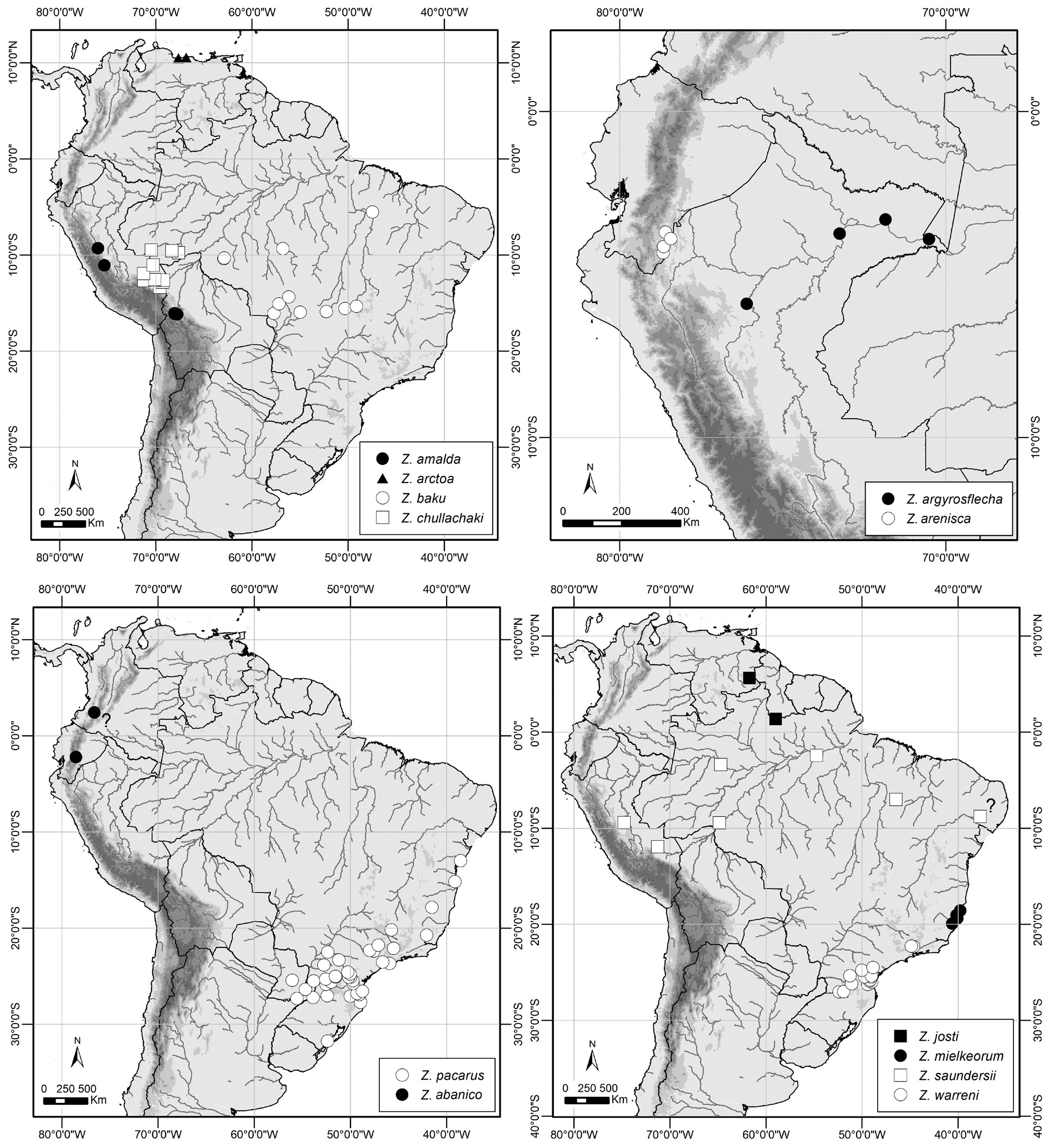
Distribution ( Fig. 14 View Fig )
This species is known to date from a highly restricted range (ca 70 km between the most northerly and southerly localities) in the Cordillera del Cóndor, an eastern outlier of the tropical Andes that lies along the border of Ecuador and Peru, and several ridges in the adjacent Río Zamora Valley in southeastern Ecuador.
Remarks
A first instar was found in one dissected female (SN-17-174) and its head capsule is illustrated in Fig. 13. A View Fig female specimen from Alfonso Ugarte, Amazonas, Peru in MUSM may represent this species, but the sclerotized portion of ductus bursae extends about three-fourths the distance from ostium bursae to corpus bursae, which contrasts with the rather short ‘sclerotized tube’ of the two barcoded females from Zamora-Chinchipe, Ecuador (MUSM-LEP-08939 and MUSM-LEP-64858; see Fig. 1 View Fig ). Based on this inconsistency, a female specimen in MUSM [Alfonso Ugarte, 2–3 km N PV3; [3°54’S, 78°26’W]; 1600–1750 m; 16 Jul. 1994; G. Lamas leg.; MUSM-LEP-105658] cannot be identified reliably as Z. arenisca sp. nov. and is, therefore, excluded from the type series.
Zischkaia arenisca sp. nov. is recovered as sister to Z. chullachaki sp. nov. based on COI barcode data (see Fig. 1 View Fig ), but the genetic distance between these two taxa is atypically low for divergence between species ( Table 4 View Table 4 ). Such low divergence might suggest conspecificity of these taxa, but the two species are recovered as moderately supported clades, with a number of consistent morphological differences (see above). In particular, the elongate, ‘egg’-shaped VHW ocelli and reduced, less prominent dorsal androconial scales of Z. arenisca sp. nov. support regarding these taxa as distinct species rather than as conspecific.
In southeastern Ecuador, this species is known from three localities, all of which support a species of climbing bamboo. Two of these sites are sandstone ‘tepuis’ that have been mined for their sand, evidently because of its purity, and the poor soils on the slopes and ridges apparently support only stunted forest. An unidentified species of Chusquea Kunth ( Poaceae : Bambusoideae ) (identified by Lynn Clark) was common at one site, the ridge east of San Roque, where it grew abundantly over remnant trees and bushes along the edges of the dirt road to the mine site on the ridge top. A similar, or the same, bamboo species was likewise common along the Sumak Yaku mine road (see Fig. 15A View Fig ), and was observed along the edges of forest and inside disturbed forest beside the village of Shaime, although it was absent along most other forest trails and roads in the region. Two females were collected at the edge of a large bamboo patch on the ridgetop at San Roque (see Fig. 15B View Fig ), flying and resting within 2 m of the ground from 09:00 to 12:30, while two other females were flying along the edge of a dirt road near bamboo from 11:30–12:30. A number of males were observed and collected flying rapidly 1–4 m above the ground, from 10:25 to 11:20, in a sunny but sheltered corridor between two patches of vegetation covered with bamboo near a ridgetop at Sumak Yaku. The presence of multiple males and repeated flight paths suggests territorial behavior. Numerous males were also collected at the same site in traps baited with rotting fish, hung 1 m above the ground in the understorey of secondary forest with abundant bamboo. Despite a reasonable amount of effort collecting and trapping at other forest sites in the region, Z. arenisca sp. nov. was not recorded anywhere else, highlighting its apparent dependence on the climbing bamboo with which the adult butterflies were always closely associated.






| FLMNH |
Florida Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |