
Myzomela wahe, Rheindt & Prawiradilaga & Ashar & Lee & Wu & Ng, 2020
|
publication ID |
https://doi.org/ 10.5281/zenodo.3608758 |
|
publication LSID |
lsid:zoobank.org:pub:8114B399-C68D-43C2-B6D3-B51AA898431E |
|
DOI |
https://doi.org/10.5281/zenodo.3610306 |
|
persistent identifier |
https://treatment.plazi.org/id/675E92C2-05D3-4C98-B861-A7A99CA7AE9E |
|
taxon LSID |
lsid:zoobank.org:act:675E92C2-05D3-4C98-B861-A7A99CA7AE9E |
|
treatment provided by |
Plazi |
|
scientific name |
Myzomela wahe |
| status |
sp. nov. |
SM4:
Myzomela wahe View in CoL , species nova
(Taliabu Myzomela ;
urn:lsid:zoobank.org:act:675E92C2-05D3-4C98-B861-A7A99CA7AE9E
) Frank E. Rheindt, Dewi M. Prawiradilaga, Hidayat Ashari, Suparno, Meng Yue Wu
Holotype
MZB.Ornit.34.357 ( fig. S6 View Fig ); adult male collected 9 Dec 2013 near Waiyo dinahana Camp (~ 1200m) on Taliabu Island (S 01⁰ 47.614 '; E 124⁰ 48.216 '). Collected by the Rheindt / LIPI field party, including tissue samples from breast muscle and liver; skin prepared by Suparno; field number Tbu21; no molt; medium fat; weight 8g; wing length 5.7cm; wing spread 16.9cm; total length 10.1cm; bill 1.9cm; tail 3.9cm; tarsus 1.8cm.
GoogleMapsDescription of holotype
Head and mantle down to uppertail coverts and to central belly are bright scarlet-red (7.5R 4/14) with subtle blackish suffusion owing to individual feathers having broad red distal half followed by black and then increasingly greyish basal half; jet-black narrow orbital ring and loral line. Upperwings, sides of mantle, scapulars and rectrices are matt black, with the remiges appearing slightly duskier black because of narrow olive edging to outer webs; underwing coverts and outer webs of the underside of remiges are whitish; a blackish-grey (N1.5) wedge extends from vent up to central belly. Iris dark-brown; bill all-black; legs grey with yellowish soles.
Diagnosis
The diagnosis refers to the adult male plumage only. Female Myzomela individuals of the Wallacean radiation are generally very similar across species, and more research is needed to identify good distinguishing marks.
The new species is an obvious, bright scarlet-and-black representative of the Wallacean grouping of Myzomela honeyeaters that have often been united within the Myzomela sanguinolenta complex (6 1, 7 5), but have been considered a radiation of independent species by more recent authors [e.g. ( 57)]. Within this radiation, the new species most closely resembles M. chloroptera from Sulawesi and M. elisabethae from Seram.
Compared to M. chloroptera , the new species has a more extensive and brighter red body and blacker wings, scapulars, and tail. Scarlet feathers in the new species are mostly black and grey at the base, with only the distal half scarlet, and with a sharp contrast between scarlet and black, lacking M. chloroptera ’s narrow yellow bands in between the red and black portions of each individual feather that render the latter’s overall body more orange-tinted. In M. chloroptera , the red is not as extensive on the underparts, and the abdomen and vent are paler buff-white versus blackish-grey in the new species.
Compared to M. elisabethae , the new species is paler and greyer (not black) on vent and abdomen. It also has a smudgier scarlet body coloration (caused by the black and grey basal halves of individual feathers), with dark smudges especially pronounced on face and on breast sides, as compared to more uniform red underparts, head and mantle in M. elisabethae . Black lores in the new species are wider and not as clearly delimited as in M. elisabethae .
Compared to M. wakoloensis from Buru, the new species differs in its all-black upperwing coverts (versus red shoulders and carpal bar), and in its blackish to brownish-grey belly and vent (versus red color reaching to vent).
The new species conspicuously differs from all other Wallacean representatives of the M. sanguinolenta radiation, such as M. boiei (Banda Islands, Tanimbar) , M. vulnerata (Timor) , M. batjanensis (Bacan, Obi) , M. dammermani (Sumba) , M. irianawidodoae [Rote; ( 73)] and M. kuehni (Wetar) , in its extensive red coloration reaching from head to belly and rump ( 57).
Etymology
We name this species after the village of Wahe (1.716°S, 124.843°E), which is the gateway to the highest elevations on Taliabu where it most commonly occurs. The species epithet is an indeclinable noun in apposition.
Individual, age-related and sexual variation within taxon
Two adult male specimens (holotype and 34.356) are virtually identical in coloration. Two additional male specimens (34.373 and 34.403), most likely immatures, are similar but have the scarlet body color replaced with a more rose-red hue (5R 4/14); they also do not exhibit any distinct olive margins of remiges. No comparative specimen material exists for adult females of the new species, which are generally similar among all regional Myzomela species. Photos confirm that the female of the new species resembles other regional species (data not shown). One juvenile specimen (34.407) lacks red coloration and is generally drab grayish-brown (paler on underparts), becoming more intense earthen-brown on the mantle, rump and upperwing coverts, with the underside of remiges and rectrices mid-grey.
History of discovery
This new Myzomela was first spotted by a University of East Anglia expedition to Taliabu in 1991 led by P.J. Davidson. Participants of this exploratory survey were probably the first ornithologists to visit the highlands of Taliabu between 600-800 m elevation (64, 9 8), where they found the new species predominantly at the highest altitude they reached (roughly 800m). FER then encountered the species commonly during his visit to the highlands of western Taliabu between 4-18 April 2009 ( 19, 48), where we returned between 6-16 December 2013 to collect the holotype and accompanying individuals ( 19).
Distribution and status
This new Myzomela has thus far only been found on the island of Taliabu within the Sula Archipelago. On Taliabu, we have recorded it from sea level to 1,300m ( 19, 48), and it presumably occurs further up all the way to the highest elevation at ~1,415m. The species inhabits forest canopy and edge habitat, and is a nectarivore and frugivore that has been photographed feeding at flowers ( 48). Akin to multiple other Myzomela species in Wallacea ( 57), the new species is seen far more often at higher elevations (roughly above ~800m) than in the lowlands. Davidson et al. ( 64), who first sighted this species, exclusively found it at higher elevations, mostly around ~800m, and we recorded it as “abundant” above 800m, whereas we have only had a handful of sightings in the lowlands. It is unknown to what extent elevational differences in abundance reflect the difficulty of detecting a canopy bird in tall lowland forest as compared to stunted montane forest. However, although detection difficulties may help mask its true abundance in the lowlands, we do not believe it is possible for the new Myzomela to be equally common across elevations given that other small canopy birds do not escape detection in the lowlands and are recorded commonly. In fact, the new Myzomela may even be a breeding endemic to montane elevations and only venture into the lowlands seasonally, following tree flowering and fruiting events. This potential seasonal displacement would explain why some expeditions have completely overlooked it in the lowlands while others have recorded it on occasion.
It is contentious whether the new Myzomela is endemic to Taliabu, or also occurs on the other two main islands of the Sula Archipelago (Mangole and Sanana). The latter two have received even less ornithological attention than Taliabu, and to the best of our knowledge, the Myzomela has never been recorded there on recent ornithological expeditions (D. Yong, pers. comm.; J.A. Eaton, pers. comm.). Sanana is relatively low-lying (up to ~600m) and therefore likely to lack a number of Taliabu’s montane species. Mangole reaches its highest peak at 1,127m, but areas above 800m are confined to ~328 ha only, which gives it a comparatively small area of true montane forest. The new Myzomela ’s level of dependence on vegetation at high elevations probably determines whether it would be capable of occurring on Mangole or even Sanana. Future fieldwork on those two islands will shed light on whether the new species occurs there or not.
FER and Filip Verbelen first discovered a new Myzomela population in the mountains of Peleng in the neighboring Banggai Archipelago during a visit between 23-30 March 2009 and recorded it as reasonably common at elevations between 700-900m in the western Peleng highlands ( 49). On our collecting expedition, we only had one aural encounter of this Myzomela on Peleng on 21 Dec 2013, indicating that the population might have been seasonally displaced or more retiring ( 19). However, photographic evidence from the 2009 visit indicates that the Peleng population has a plumage coloration that is very similar – if not identical – to M. chloroptera from Sulawesi and quite different from M. wahe . As this Peleng population remains uncollected, future fieldwork will hopefully produce specimens allowing a determination whether there are differences between the Peleng birds and M. chloroptera .
Taxonomic rationale
The new Myzomela from Taliabu is obviously a member of the M. sanguinolenta radiation, which used to be considered a single wide-ranging polytypic species for a long time (6 1, 7 5), but has now been divided into multiple monotypic island endemics by most recent authors on the basis of extensive plumage differences [e.g. (57, 8 0, 99)]. Within this radiation, its plumage most closely resembles that of M. chloroptera from Sulawesi and M. elisabethae from Seram, but is discrete from both of them in multiple characters (see Diagnosis). Its morphological differences from both M. chloroptera and M. elisabethae exceed those between other closely related species pairs within the M. sanguinolenta radiation, such as M. dammermani and M. irianawidodoae ( 73). Therefore, its diagnosis here as a species level taxon rests on Mayr and Ashlock’s ( 70) yardstick approach, by which species status is conferred on the basis of divergences that exceed those of species pairs in which biologically important differences have been unequivocally demonstrated.
Myzomela honeyeaters have a wide vocal repertoire that is oftentimes problematic in the context of bioacoustic analysis ( 73). The Taliabu Myzomela was hardly vocal during our encounters, and we do not consider vocalizations a fruitful taxonomic avenue in delimiting this new species from others. Instead, we have generated mitochondrial sequence data to further shed light on the new species’ taxonomic status.
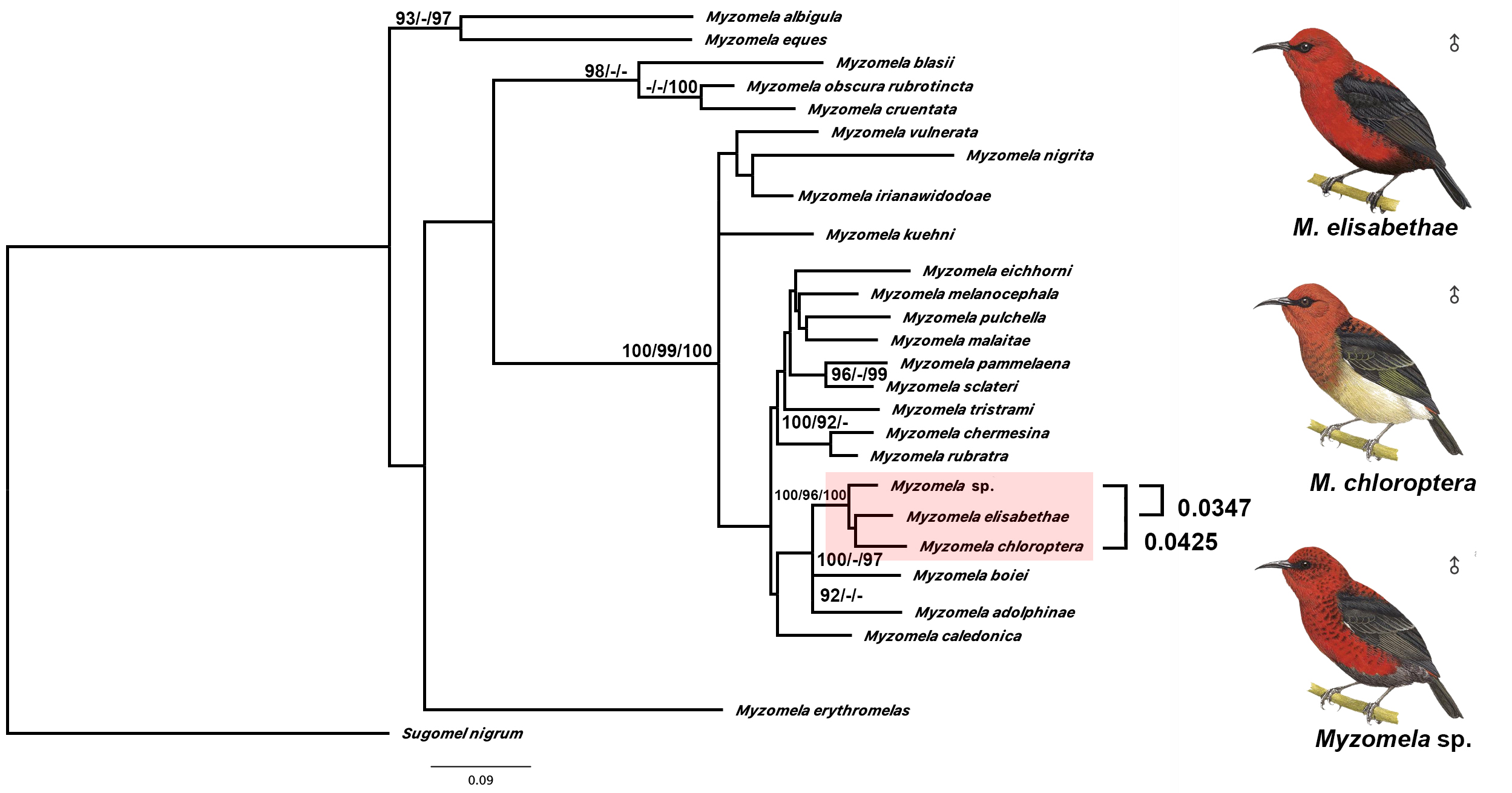
Mitochondrial evidence: Our mitochondrial tree analysis was based on sequences of the coding gene NADH dehydrogenase subunit 2 (ND2), comparing M. wahe to a great majority of members of the eastern and central Indonesian radiation of Myzomela honeyeaters. Results indicated that M. wahe is most closely related to geographically adjacent species such as M. chloroptera from Sulawesi and M. elisabethae from Seram ( fig. S7 View Fig ). Our analysis does not provide resolution on whether either M. chloroptera or M. elisabethae constitute the sister species to M. wahe , and in fact, the latter two may well be sister to each other instead ( fig. S7 View Fig ), rendering M. wahe a more basal species. M. wakoloensis from adjacent Buru is not represented in our mitochondrial dataset, and likely forms part of this group. Based on plumage coloration, M. wakoloensis is frequently considered to constitute the sister species of M. elisabethae , and the two are sometimes merged under one species [e.g. ( 99)]. Therefore, we assume that M. wakoloensis – albeit closely related – will probably not emerge as the direct sister species of M. wahe in future analyses.
Mitochondrial ND2 divergence values between M. wahe and its two most closely related relatives ( M. elisabethae , M. chloroptera ) range between 3.5%–4.3% ( fig. S7 View Fig ), well above the mitochondrial divergence threshold of 2-3% widely postulated in the literature (10 0, 10 1), providing additional support for species level recognition of M. wahe .
Methodology
Mitochondrial analysis: We extracted DNA from the breast muscle tissue of all our Myzomela wahe material (n = 5), and from four independently collected Myzomela irianawidodoae samples ( 73), using the Qiagen DNeasy Blood and Tissue Kit (Qiagen, Germany) following the manufacturer’s instructions. ND2 was then sequenced following Lim et al. (10 2).
Using CodonCode Aligner version 8.0.2. (CodonCode, USA), our ND2 sequences were trimmed and assembled. These sequences of 991 bp length were then aligned in MEGA7 (9 5) with the ND2 sequences of 23 other species of Myzomela generated by Marki et al. (10 3), including a sequence of Sugomel nigrum from Driskell and Christidis (10 4) as the outgroup based on its relative completeness in comparison to other potential outgroup sequences. See table S6 for information on sequences and accession numbers. We ran maximum parsimony and neighbor-joining trees with MEGA7, and computed uncorrected pairwise divergences among M. chloroptera , M. wahe and M. elisabethae . Maximum likelihood analysis was carried out in raxmlGUI (96 under the GTR + Inverse + Gamma model as identified by jModelTest 2.1.7 (97, 10 5).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |