
Tonkinomys daovantieni, Musser & Lunde & Son, 2006
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2006)3517[1:DOANGA]2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03F37B6F-DE13-9E56-28D6-6672FE85FA79 |
|
treatment provided by |
Carolina |
|
scientific name |
Tonkinomys daovantieni |
| status |
sp. nov. |
Tonkinomys daovantieni View in CoL , new species
HOLOTYPE: AMNH 275618 View Materials , an adult female collected by the authors ( DPL 1706 ) on 17 April , 2004, in the vicinity of Lân Ðăt Village (21 ° 409520W/106 ° 209280E), 150 m, Hu ,ũ Liên Nature Reserve , Hu ,ũ Liên Commune , Hu ,ũ Luñg District , Lang So ,n Province , Vietnam. The specimen was fixed in 10% formalin for several days before being transferred to 70% ethanol for storage. A liver sample was collected prior to fixation and placed in lysis buffer before its ultimate transfer to the American Museum Cryo Collection ( AMCC 125060 View Materials ). The skull ( figs. 10 View Fig , 11 View Fig ) was later extracted and cleaned by dermestids in a colony at the American Museum.
REFERRED MATERIAL: In addition to the holotype, 13 additional adults of T. daovantieni were collected from the vicinity of Lân Ðăt Village during April, 2004, on the days indicated ( AMCC catalog numbers tied to liver samples are in brackets). April 13: AMNH 275575 View Materials [125016], male; 275576 [125017], male; 275577 [125018], male. April 14: 275586 [125027], female; 275593 [125034], male. April 16: 275602 [125044], male. April 19: 275627 [125069], male; 275644 [125086], male. April 23: 275688 [125130], male. April 24: 275692 [125134], male; 275693 [125135], ear with attached bit of furry skin only. April 25: 275711 [125153], male; 275712 [125154], female .
GEOGRAPHIC DISTRIBUTION: Tonkinomys daovantieni is currently known only from the type locality, but the species probably occurs in suitable forested habitats throughout the karst landscapes of northern Vietnam (identified as the ‘‘Vietbac Karst Zone’’ by Tuyet, 1998: 187; or the Caobang and Bacson regions by Głazek, 1966), and possibly in southern China (see the distributions of karst outcrops mapped in Tuyet, 1998, and Zhang, 1989).
DIAGNOSIS: Because daovantieni is the only known species of Tonkinomys , generic and specific diagnoses are the same.
ETYMOLOGY: We are honored to name this distinctive gray-furred and short-tailed limestone endemic after the late Ðào Văn Tién, who when he died on May 3, 1995, at the age of 78, was emeritus professor of biology at the National University of Hanoi. Our esteem for this Vietnamese scholar is reflected in the obituary prepared by Groves and Weitzel (1995: 15216) :
Professor Tien was educated in Hanoi under the French colonial Administration. He taught several generations of Vietnamese scientists and carried his Biology Faculty through the difficult years after his country gained its freedom. Though Professor Tien’s principal works and many of his papers were in mammalian zoology, his advice, his academic research, and most importantly his student support was never limited to this. Certainly, all senior biologists in northern Vietnam today gladly call Professor Tien their teacher. It is hard to calculate the importance of this gentle and thoughtful man in the history of Vietnam’s science. He was the father of his field in Vietnam. His loss was a great blow for us, though he left for the world an incalculable legacy of studies of the rich biology of Vietnam, and a scholarly tradition for those who will follow in his footsteps.
The biodiversity surveys undertaken in northern Vietnam during the past decade that have contributed so significantly to knowledge of mammalian diversity in that country can be viewed as extensions of earlier vertebrate surveys in northern Vietnam from 1957 to 1971 initiated and managed by Professor Tién. His results generated large and important collections of voucher specimens (currently housed at the University of Hanoi), and were summarized by him in 1985 ( Ðào, 1985). That body of field data and museum specimens, and the scientific results derived from them formed the foundation of ‘‘Checklist of Mammals in Vietnam’’ published nine years later ( Ðăng et al., 1994), which at the time stood as the finest and most comprehensive annotated list of Vietnam mammals, and today remains not only an important reference, but a tribute to Professor Tién, his students, and his colleagues.
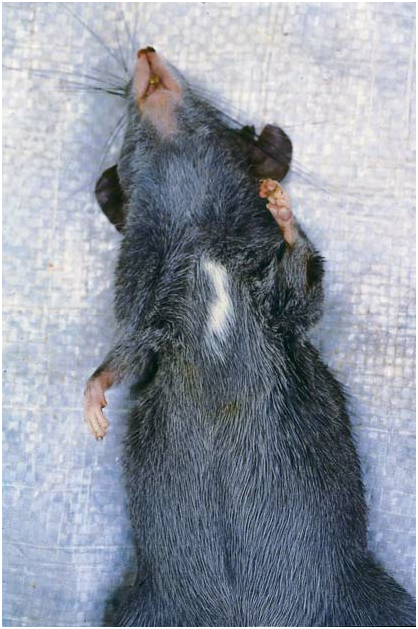
MORPHOLOGICAL DESCRIPTION: Large head and eyes, a stocky body, dark gray ears, short and bicolored tail, grayish black (gunmetal) upperparts, and dark gray underparts charac- terize live Tonkinomys daovantieni ( figs. 4 View Fig , 5 View Fig ; table 1). Detailed descriptions of its external features, skull, and dentition are provided below.
External traits: The dense fur covering upperparts of head and body is grayish black with burnished highlights, and consists of three types of hairs. The soft underhairs (up to 12 mm long) are gray for their entire length. The overfur layer (up to 15 mm thick) is formed by wide, flexible, and grooved spines, each pale gray for most of its length and tipped with brown or black. Scattered guard hairs project 5210 mm beyond the overhairs, the basal half of each is gray, the distal half black. The combination of hair types and their pigmented banding patterns imparts a thick and semispinous texture, and a burnished, grayish black tone to the upperparts. The dorsal pelage is less spiny along the sides of the body, appears especially soft and gray over the shoulders and hips, and gradually blends into the softer and shorter (up to 8 mm thick) dark gray fur covering the underparts. That coat is formed by soft underhairs mixed with soft, wide, and grooved spines; both kinds of hairs are gray for their entire lengths. Although paler than the dorsal coat because it has no dark brown guard hairs, no sharp demarcation exists between dorsal and ventral fur. A white patch (size ranges from 10 mm long and 5 mm wide to 30 mm by 12 mm) interrupts the dark gray fur over the chest in 11 of the 13 specimens; the other two lack this chest mark. On the forehead a blaze of white hairs (expression ranges from a few hairs to a patch 15 mm long and 5 mm wide) contrasting with surrounding dark fur is present in nine specimens; four, including the holotype, lack the blaze. Sides of the face and throat are dark gray; the lips, chin, and rhinarium are unpigmented. Fur covering the lower limbs is dark gray everywhere, but shorter than that on the torso.
Eyes, relative to head area, are large, proportionally similar to species of Rattus . A wide unpigmented ring surrounds each narrow and circular, dark brown eyelid. Beyond the eyes, the fur is dark gray covering this part of the head, and although somewhat more suffused with black, this darker tone does not define a facial mask.
The pinnae (external ears) are moderately large, but not disproportionately expansive relative to body size. They are dark gray, somewhat oval in outline, and rubbery in texture. Short brown hairs (visible only under magnification) are sparsely scattered over both outer and inner surfaces, but do not tint the gray background.
The mystacial vibrissae are either dark brown or unpigmented, fine, and very long (up to 75 mm); when laid against the head the longest projects up to 20 mm beyond the distal margin of each pinna. The longest in each pair of superciliary (or supraorbital) vibrissae (up to 60 mm long) also extend well past the pinna when appressed against the head. The short submental and interramal vibrissae are unpigmented. The few short (up to 25 mm) genal vibrissae are inconspicuous and unpigmented for most of their lengths. The base of the wrist supports four moderately long (up to 10 mm) unpigmented ulnar carpal vibrissae, and from the inside of each heel springs a longer (up to 15 mm) unpigmented tarsal vibrissa.
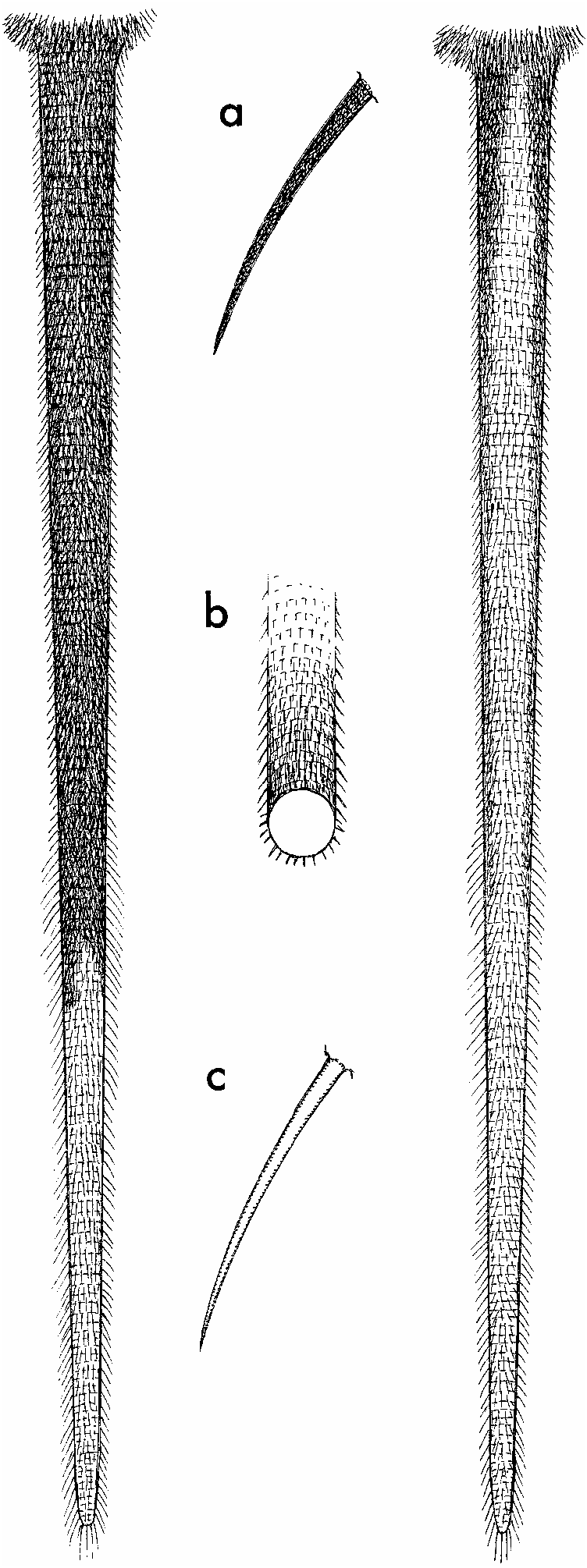
The tail is appreciably shorter than the length of head and body (LT/LHB 5 83%, average of 13 individuals) and it is thick (range in 13 specimens is 5.7 mm to 6.3 mm, measured laterally at midpoint of tail length) and round in cross section (table 1; fig. 6 View Fig ). The proximal one-half to three-fourths is dark brown on dorsal and lateral surfaces (produced by the brown pigment in scales and scale hairs). The ventral surface is white, as are all surfaces of the distal one-fourth to one-half (25%–44% of total tail length among the 13 specimens). In a few specimens, scattered scales in the unpigmented regions retain very pale brown pigment, which provides a slight speckling to the white portion, usually over the ventral surface. Scales in the overlapping annuli are moderately large (10 annuli of scales per centimeter, counted along basal one-third of the tail), and beneath each scale emerge three hairs. These are bristle-like and about as long as two scale annuli (1.0 mm) near the base of the tail, but those at the tail tip are softer and longer (526 mm); there is a gradual increase in hair length and texture from base to tail tip. Because the hairs are mostly longer than the scale annuli, the tail appears somewhat hairy, especially the distal half ( fig. 6 View Fig ).
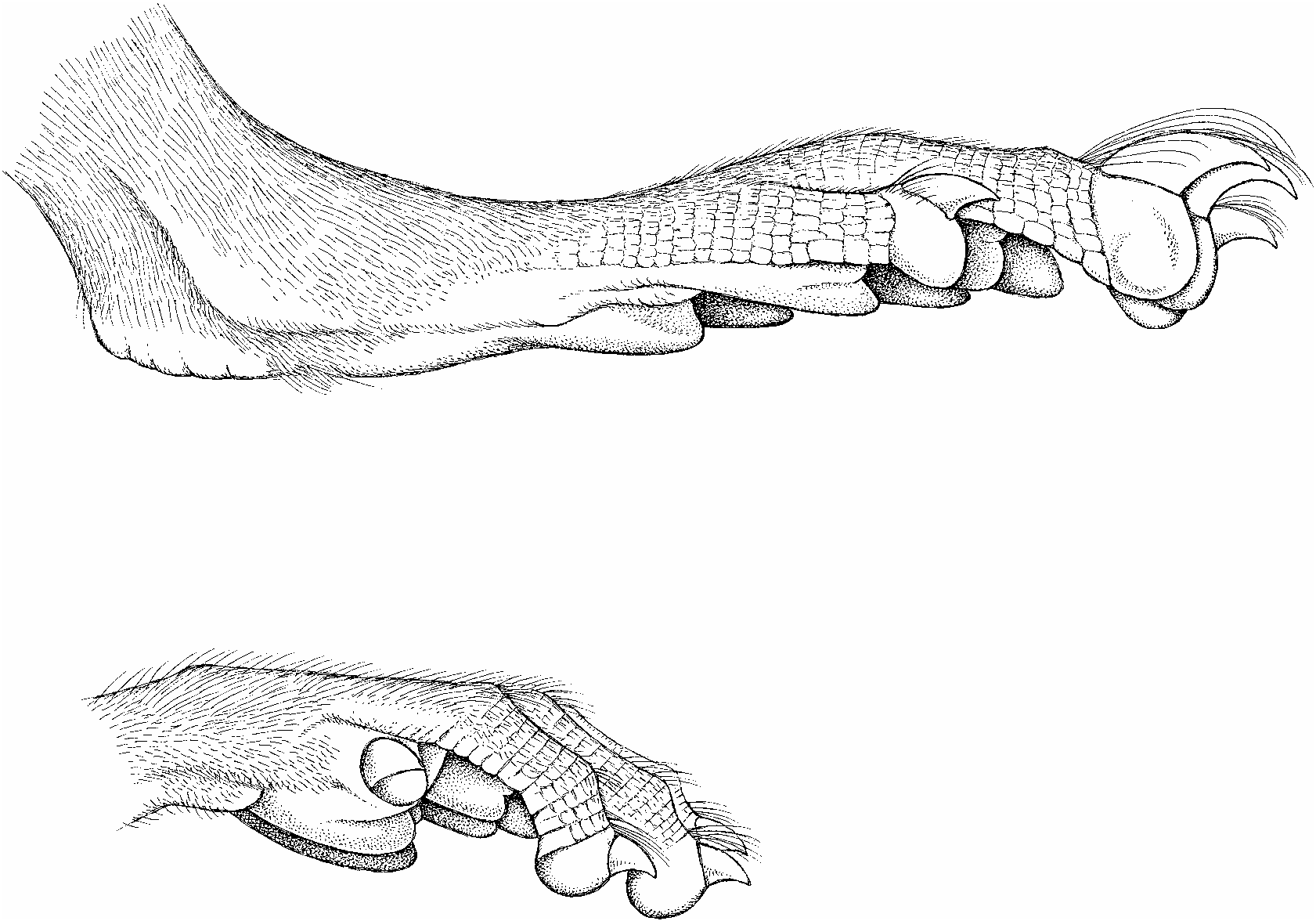
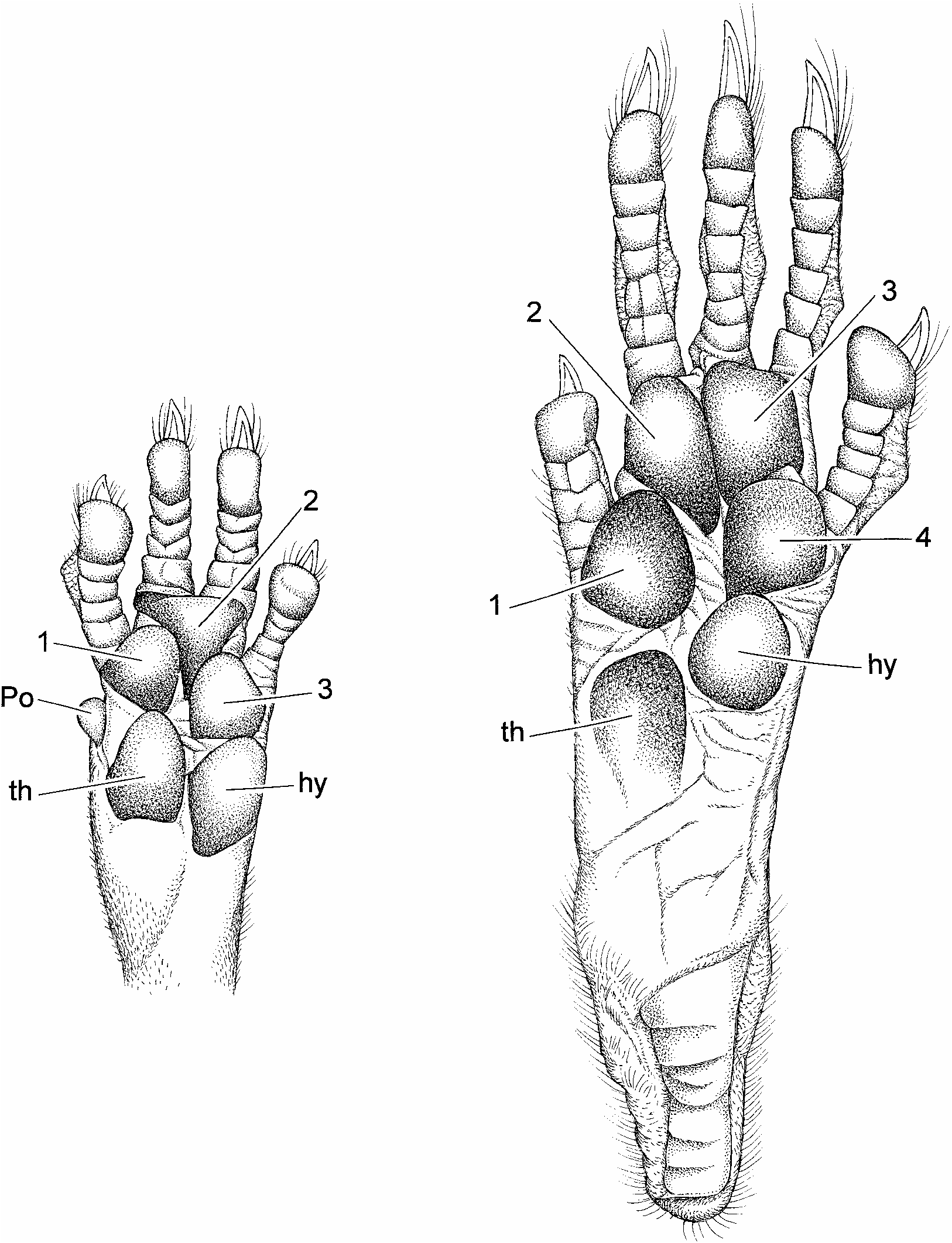
Claws are unpigmented, as are their thin covering of hairs (ungual tufts) and the digits of the front and hind feet. So too is the dorsal metacarpal and metatarsal integument, but an overlying covering of brown hairs imparts a diluted brown wash to these surfaces. Both front and hind claws are not unusually long, and are gently curved ( fig. 7 View Fig ). Their covering ungual tufts either do not or only slightly project beyond the claw tips ( fig. 8 View Fig ). Each palmar surface is naked and mostly unpigmented. Its topography is dominated by five huge and swollen pads: three interdigital, thenar, and hypothenar ( fig. 8 View Fig ). Gray patches mark some of the pads, and the ventral surfaces of all are nearly smooth, exhibiting only traces of semicircular and transverse lamellae. The hind foot is moderately long and wide (table 1). Its naked plantar surface and some of the pads are dark gray. The anterior half of the plantar surface is formed by six very large and swollen pads (tubercles) in a tight cluster: two inner interdigitals (2 and 3), two outer interdigitals nearly opposite each other (1 and 4), a large hypothenar (nearly as large as an interdigital) just behind and slightly overlapping the lateral interdigital, and an even larger thenar ( fig. 8 View Fig ). Size and protuberance of the pads relative to plantar surface, and their contiguous positions resemble the conformation seen in arboreal murines like the Sulawesian species of Margaretamys ( Musser, 1981: 279) . They contrast with such terrestrial examples as Sundanese Sundamys muelleri and Berlymys bowersii ( Musser and Newcomb, 1983: 357), Rattus argentiventer ( Musser, 1973: 7) , or Sulawesian Maxomys ( Musser 1991: 27) whose pads are smaller relative to area of the plantar surface with the two outer interdigital mounds set well apart from the inner pair, and the hypothenar and thenar widely spaced from each other and the interdigitals.
Each female in our sample has four pairs of mammae: one pectoral, one postaxillary, and two inguinal.
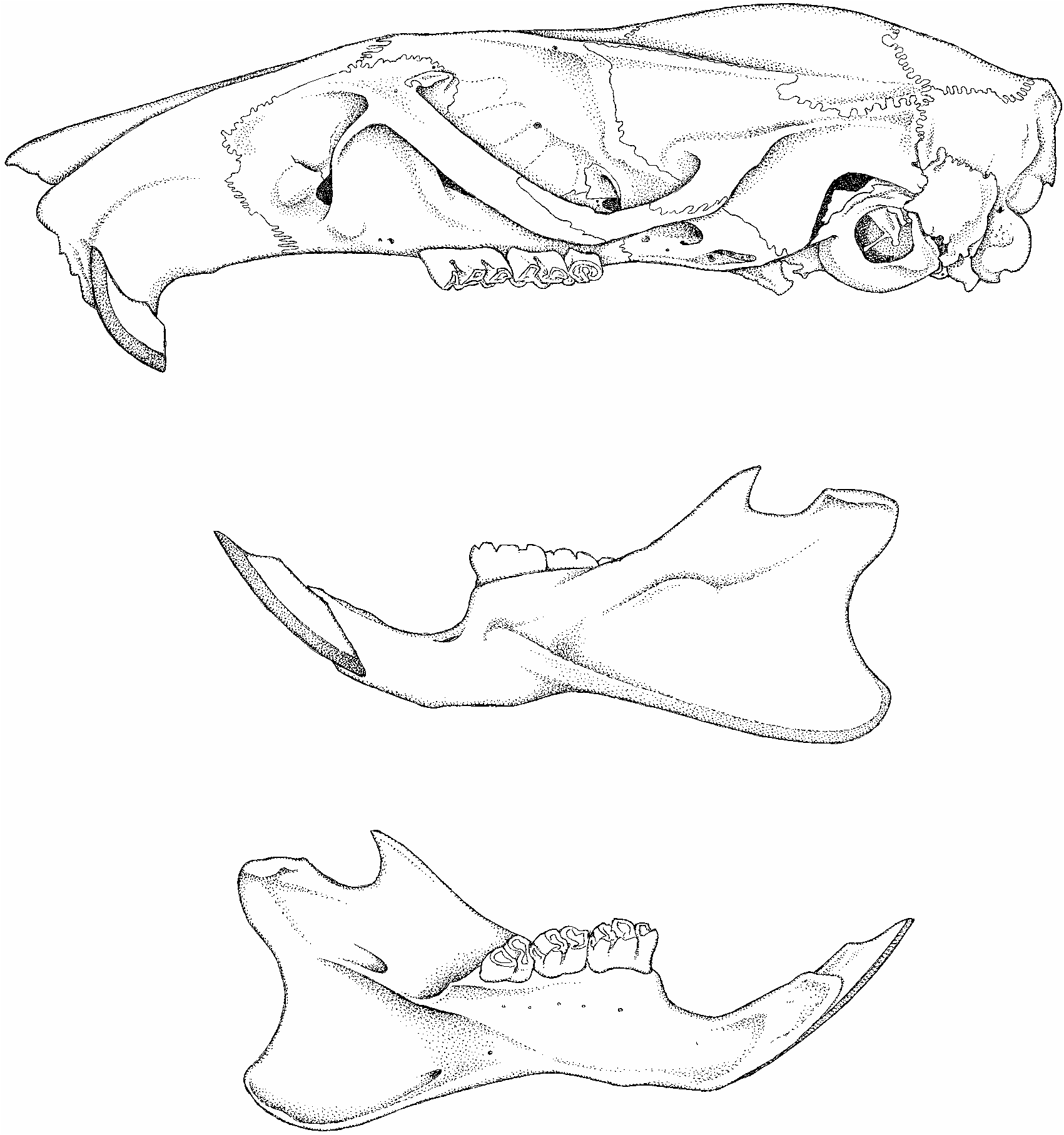
Skull: Tonkinomys daovantieni has an elongate skull, sturdy in appearance, the visual impression reflecting a long rostrum, narrow cranium, and robust zygoma. In dorsal view ( fig. 10 View Fig ), the rostrum is moderately long and wide, its parallel sides dimpled by a small nasolacrimal capsule (just anterior to the anterior spine of each zygomatic plate). The distal portion of the rostrum is slightly tapered, the anterior margin of the nasals is triangular, and the adjacent premaxillaries project well beyond the posterior nasal-frontal suture. A moderately deep zygomatic notch is evident between the rostral wall and the projecting anterior spine of the zygomatic plate. The dorsal portion of each lacrimal bone (in the anterolateral pocket of the orbit) is small, rectangular, and fused with the dorsal maxillary zygomatic root (no shared suture with the frontal). Dorsolateral margins of the interorbit, which is about as wide as the rostrum, are defined by high ridges that are prominent as they sweep back along the dorsolateral postorbital margins of the frontals; these ridges become weaker in expression and less pronounced where they extend (as temporal ridges) onto the parietal margins (accentuating dorsolateral rims of the cranium) to about the level of the posterior edge of the squamosal zygomatic root. Here the temporal ridge is barely discernable, and the parietal extends ventrally to form the dorsolateral cranial wall between the posterior margin of the zygomatic root and the lamboidal ridge; a bony swelling outlines the parietal-squamosal connection all the way to the lamboidal ridge. The interparietal is long (in anterior-posterior plane) and moderately wide (its lateral margins are within the parietal and supraoccipital sutures, not extending to the parietal-squamosal suture) and its posterior two-thirds roofs a deep occiput with a slightly convex or chevronshaped posterior margin. Zygomatic arches are robust and arranged nearly parallel to one another; neither zygoma bows out far from the skull.
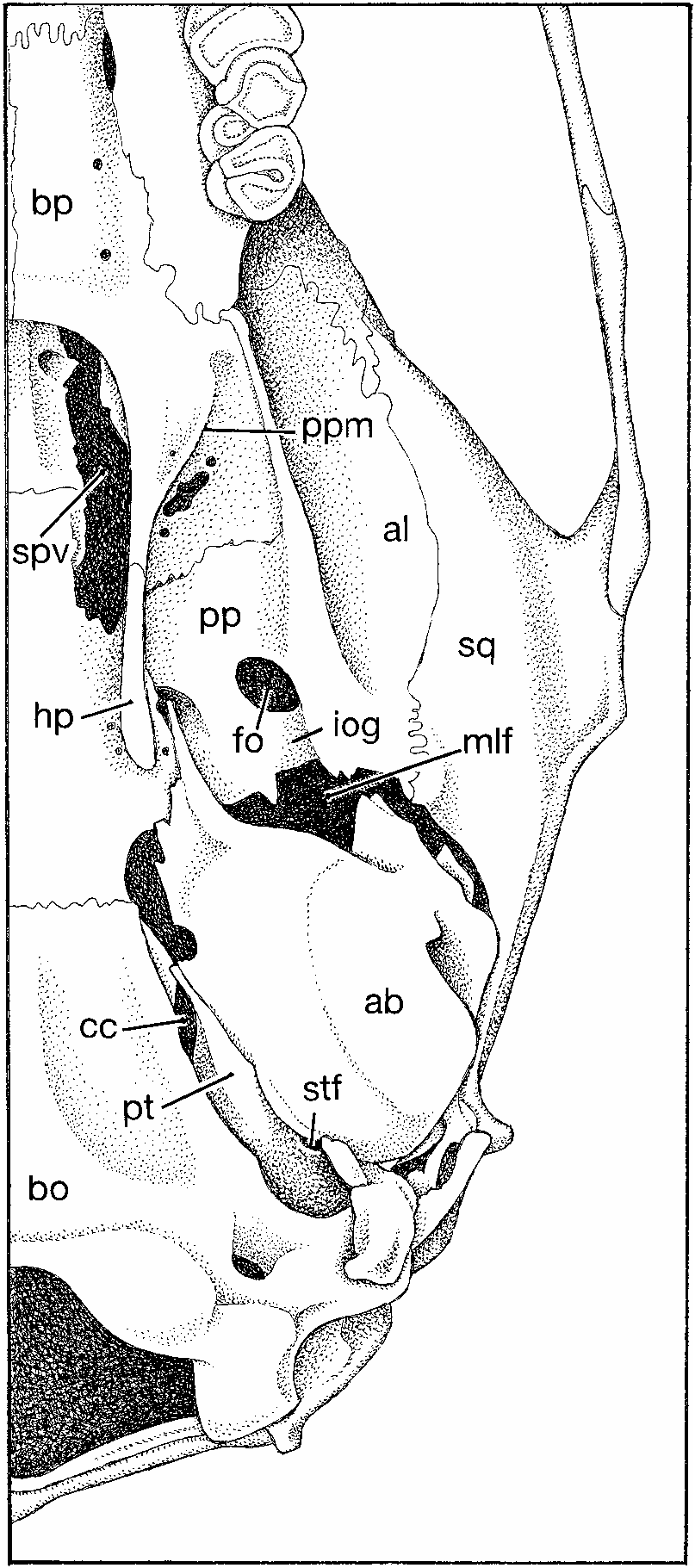
The skull appears long, low, and without significant basicranial flexion when viewed from lateral perspective ( fig. 11 View Fig ). The rostrum is rectangular in side view, and extends slightly beyond the incisor faces to enclose the nasal aperature in a short tube in which nasals provide the roof and premaxillaries the sides. Much of the ventral maxillary root of each zygomatic arch lies anterior to the molar row, with the posterior margin of the zygomatic plate extending behind the front margin of the toothrow (0.521.0 mm is the range in our sample of skulls). The zygomatic plate is wide, its convex anterior margin projecting beyond the dorsal maxillary zygomatic root to form a moderately deep zygomatic notch (best seen in dorsal view). This anterior spine of the plate does not project far enough forward to conceal the nasolacrimal capsule, which is small and only slightly inflated. Maxillary and squamosal zygomatic processes are joined by a long jugal to form a thick and robust zygomatic arch. The squamosal root of each arch is situated high on the side of the cranium where its posterior ridge2like portion extends horizontally just below the posteroventral margin of the parietal, which projects ventrad to form the dorsolateral side of the cranium. The low ridge of the squamosal zygomatic root disappears 10–15 mm anterior to the vertical ridge formed by the squamosal-exoccipital suture. The configuration is similar to, but not identical with, that documented for species of Leopoldamys (see Musser, 1981:fig. 18) and Niviventer (Musser et al., 2005: figs. 7 View Fig and 8 View Fig ). Dorsal to the auditory (ectotympanic) bulla and posterior to the squamosal zygomatic root, the squamosal forming the side of the cranium is intact except for a minute subsquamosal foramen at the prominent ridge formed by the squamosal-exoccipital suture. This foramen is the only landmark identifying where the squamosal and a wide hamular process of the squamosal have completely fused to form the solid cranial wall. A spacious postglenoid foramen that is confluent in some specimens with a wide postalar fissure (ventral to the postglenoid foramen) separates the squamosal margin from the periotic flange and dorsoanterior surface of the ectotympanic (auditory) bulla. That capsule is attached to the squamosal dorsally by the posteroventral portion of that bone, which represents the cranial wall fused with a wide hamular process. There is no anterior attachment, which is the usual pattern among most murines. An anterior flange of the periotic, the tegmen tympani, is present (and partly separates the dorsal postglenoid foramen from the ventral postalar fissure), but is separated by a narrow or moderately wide gap from the posterior margin of the squamosal in most specimens; there is no posterior suspensory process of the squamosal to which the tegmen tympani could attach (in some cricetid rodents, the tegmen tympani overlaps a posterior projection of the squamosal to form one of the suspensory attachments between ectotympanic capsule and squamosal; see the diagrams in Voss, 1993: 19). The moderately inflated mastoid is tightly sutured to squamosal and exoccipital. It is completely ossified, without the moderate or large fenestrae found in some murines ( Crunomys celebensis , for example; Musser, 1982: 38) and sigmodontines (see Carleton and Musser, 1989: 34); only a very small opening penetrates the dorsal mastoid-exocci- pital suture. The foramen magnum is directed caudad (which reflects the horizontal rather than arched axis of the skull—no significant basicranial flexion), and the deep occiput projects slightly beyond posterior surfaces of the occipital condyles. The ethmoid foramen within the orbit is tiny, and the optic foramen moderately large. The dorsal palatine foramen is separate from the sphenopalatine foramen, a configuration usual among Asian murines, with a few exceptions where the two foramina are coalesced ( Musser, 1982: 22). Eight specimens have a bony alisphenoid strut (ranging from moderately wide to slender in the sample) on each side of the skull posterior to the orbit and dorsad to the pterygoid ridge (the lateral edge of the pterygoid plate). The strut separates the foramen ovale accessorius from the combined buccinator-masticatory foramen on the side of the cranium, and obscures the anterior opening of the alisphenoid canal. One specimen exhibits a strong strut on the left side, but only a bony thread on the right. Only one individual in our sample of skulls lacks struts (which are represented by only a dorsal nubbin on each side). In this specimen, the opening to the alisphenoid canal is evident, the foramen ovale accessorius and buccinatormasticatory foramina coalesce (basically disappear), and the course of the buccinator and masticatory nerves is marked by a shallow groove extending from the foramen ovale onto the side of the alisphenoid.
The short rostral tube anterior to faces of the incisors, wide rostrum, long incisive foramina and bony palate, diverging molar rows, moderately wide mesopterygoid fossa and pterygoid plates, and small bullae are the topographic highlights when the skull is studied from ventral perspective ( figs. 10 View Fig , 12 View Fig ). Just behind the incisors is a tiny interpremaxillary foramen, and beyond it the rostrum is perforated by long and spacious incisive foramina, their posterior rims situated slightly before (0.5 mm) anterior margins of the first molars, even with those molar surfaces, or projecting slightly (not more than 0.5 mm) between them. The bony palate is wide and long, extending 1.5–2.0 mm beyond the molar rows to form a broad platform with a slightly concave and smooth posterior margin (up to 20% of the palate extends past the third molars) unmarked by any bony ridge in most specimens, slightly thickened in others. Each posterolateral margin of this shelf extends as a flat, bony surface onto the anterior portion of each pterygoid plate where its sharp diagonal border marks the anteromedial margin of each pterygoid fossa. The ventral surface of the bony palate is smooth except for a pair of oblong posterior palatine foramina (in the maxillary-palatine suture opposite the middle or posterior half of each second molar), a pair of shallow grooves, and small or minute nutrient foramina scattered over the palatal shelf. The mesopterygoid fossa is wide, and dorsolateral margins of its roof are perforated by long and moderately wide sphenopalatine vacuities. On each side of the pterygoid fossa, a robust hamular process of the pterygoid marks the boundary between the mesopterygoid fossa and each adjacent pterygoid plate, its ventral surface sculptured by a shallow pterygoid fossa. The bony pterygoid plate is breached by the large ventral opening of the foramen ovale in its posterior half; its anteromedial portion along the rim of the bony palatal extension is slightly fenestrated (irregular traces of the sphenopterygoid vacuity). A shallow trough (in which the infraorbital artery passes) scores the ventral margin of the pterygoid plate between the ventral opening of the foramen ovale and the middle lacerate foramen. Small relative to overall size of the skull, the globular and only slightly inflated ectotympanic (auditory) bulla does not conceal nearly all of the periotic as it does in some other murine groups, species of Rattus , for example. In Tonkinomys daovantieni , a wide, wedge-shaped segment of the periotic is exposed between the dorsal capsular margin and basioccipital, and this wedge of periotic extends anteriorly to form the dorsal and slanting posterodorsal wall of the carotid canal. The conformation resembles that illustrated by Carleton and Musser (1989: 33) for the sigmodontine, Oligoryzomys , and is unlike many other muroid rodents in which a more inflated ectotympanic capsule covers most of the periotic and the anteromedial margin of this capsule meets the basioccipital, so the opening to the carotid canal is bounded by the ectotympanic capsule and adjacent basioccipital (the condition in Oryzomys palustris depicted by Carleton and Musser, 1989: 33; and species of Rattus and its allies illustrated in Musser and Heaney, 1992: 99). Tapering rostrally from the ectotympanic capsule is a moderately short and wide bony eustachian tube. A spacious middle lacerate foramen separates the anterior slope of the ectotympanic capsule from posterior margin of the pterygoid plate.
The cephalic arterial pattern possessed by Tonkinomys daovantieni is not unique among members of Murinae (as that subfamily is delineated by Carleton and Musser, 1984; Musser and Carleton, 2005). As in all described species of Niviventer , Chiromyscus , Saxatilomys , and Leopoldamys , T. daovantieni lacks a sphenofrontal foramen and related squamosoalisphenoid groove, the petrotympanic fissure between bullar capsule and periotic is perforated by a spacious stapedial foramen continuous laterally with a deep groove on dorsal surface of the periotic, and there is a shallow but wide groove on the posterior ventral surface of the pterygoid plate ( fig. 12 View Fig ). This set of osteological landmarks signals a cephalic arterial pattern in which the supraorbital branch of the stapedial artery is absent and the orbital circulation derives from the distal portion of the infraorbital branch. The infraorbital is a continuation of the stapedial artery, which passes over the dorsal groove in the periotic, through the middle lacerate foramen, continues in the shallow groove ventral to the posterior margin of the pterygoid plate to course through the alisphenoid canal, and emerges onto the orbital floor through the anterior alar fissure. Common to most murines ( Carleton and Musser, 1984; Musser and Newcomb, 1983; Musser and Heaney, 1992), this pattern of vessels and associated osteological landmarks is also characteristic of some sigmodontine rodents (described and diagramed for Oligoryzomys by Carleton and Musser, 1989; pattern 2 in Voss, 1988), and is derived compared with the cephalic arterial pattern hypothesized to be primitive for muroid rodents ( Bugge, 1985; Carleton, 1980).
Mandible: The sturdy dentary supports a delicate coronoid process, relative to extent of the deep ascending ramus, a moderately deep sigmoid notch between coronoid and prominent condyloid processes, and a shallowly concave posterior margin connecting condyloid and angular processes ( fig. 11 View Fig ). In contrast with the wide and conspicuous ventral masseteric ridge, the dorsal ridge is either undetectable or indicated by a shallow, elongate depression. Besides the ventral masseteric ridge, the only other significant structure that marks an otherwise smooth lateral dentary surface is a slight bulge below the anterior root of the coronoid process that in turn marks termination of the incisor alveolus within the bone. In no specimen did the incisor capsule extend farther caudad to project into the condyloid process (checked by shining a high-intensity light through the dentary) as it does in some other murines ( Musser and Heaney, 1992: 84). Topography of the lingual dentary surface resembles that characteristic of most other murines, the primary landmarks being a prominent lingual ridge above which is the large mandibular foramen (as in Papagomys armandvillei , for example; see Musser et al., 1986: 5). The body of the dentary projects anteriorly beyond the molar row as a moderately short and stocky tube containing the incisor. Just in front of the molar row on the dorsolateral labial surface is a small mental foramen.
Dentition: Enamel faces of upper and lower incisors are pale orange and smooth, without grooves or shallow sulci. Thickness of the enamel relative to the dentine is similar to that in species of Rattus and many other murines ( Musser and Heaney, 1992: 79). The upper incisors curve caudad after emerging from the rostrum (opisthodont conformation; see Thomas, 1919; Hershkovitz, 1962: 103). Gnawing edges of the uppers are at right angles to the long axis of the skull, so their combined tips form a straight cutting edge (as in species of Rattus ; see Musser and Heaney, 1992: 79), not curved or V-shaped (as in some Philippine and Sulawesi endemics; see Musser and Heaney, 1992; Musser and Durden, 2002). The lower incisors are moderately long and curved, their cutting tips either straight or arcuate.
Alveolar patterns for roots of upper molars are distinctive. A large anterior root, two robust lingual roots, and a large posterior holdfast anchor each first upper molar. No labial root is present between the large anterior and posterior anchors. Three roots hold the second and third molars in place: a large anterior and posterior, and small lingual. Two large roots anchor each lower molar. Except for the first upper molars, the pattern of roots in all our examples of Tonkinomys daovantieni is primitive for murines ( Musser and Newcomb, 1983; Musser and Heaney, 1992).
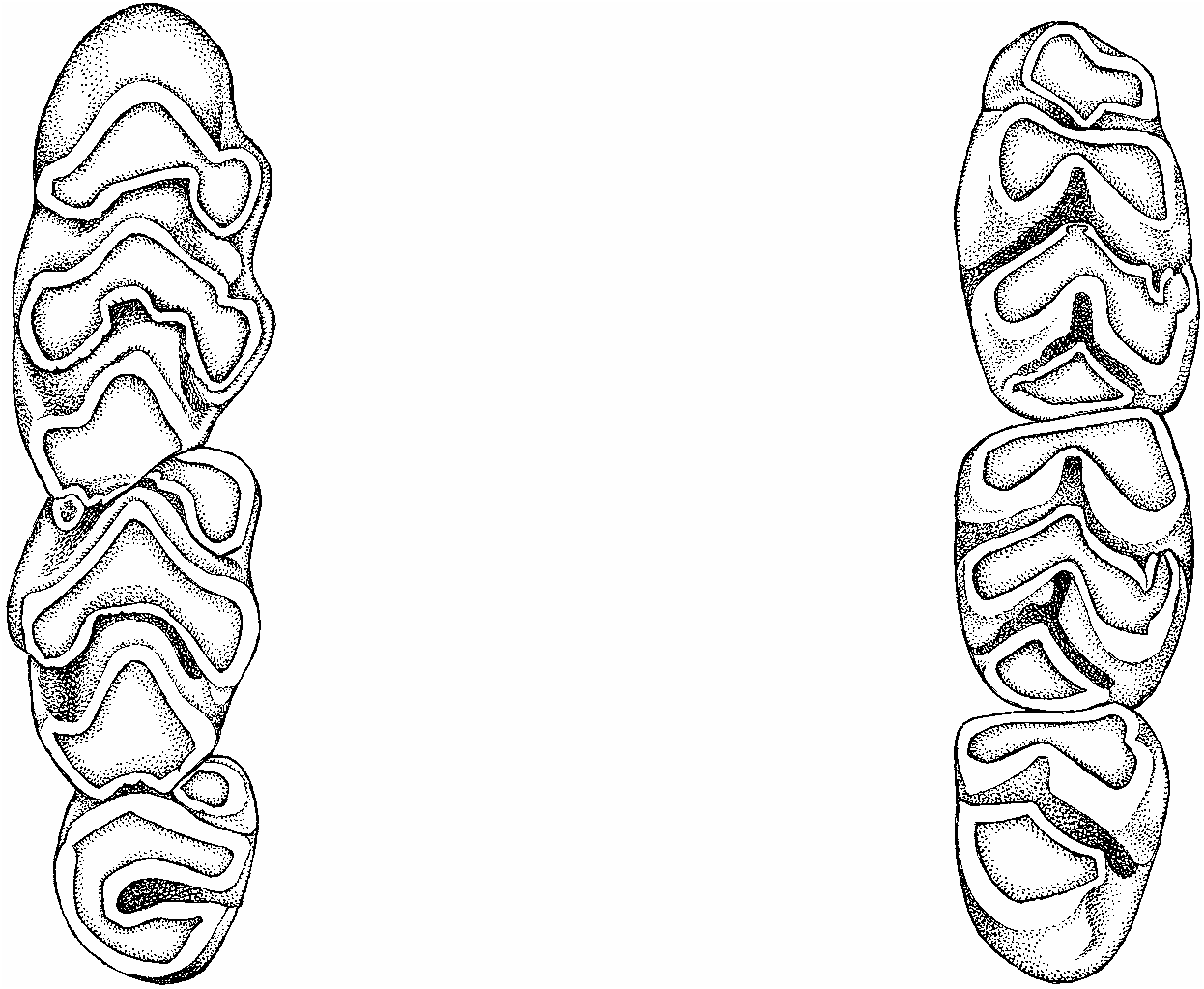
Upper molar rows are moderately long (about 17% of occipitonasal length) and strongly diverge posteriorly (width of bony palate is considerably greater between third molars than between the first; see fig. 10 View Fig ). Occlusal surfaces of the brachydont (lowcrowned) molars are simple in topography, consisting of tightly pressed rows exposing chunky chewing surfaces in the shapes of narrow chevrons and diamonds ( fig. 14 View Fig ). Occlusal surface of each first upper molar consists of two anterior rows of cusps, each in the form of a tight chevron, and a large roughly diamond-shaped posterior surface. The most anterior chevron is comprised of cusps t1, t2, and t3; cusps t2 and t3 are so coalesced that their limits are obliterated, at least in adults. The blended cusps t4, t5, and t6 form the second chevron, although the labial and lingual cusps retain some definition. The large, chunky posterior third of the molar consists primarily of cusp t8 merged with a smaller, indistinct cusp t9 to form a roughly diamond-shaped chewing surface. On the occlusal surface of each second upper molar, the anterior chevron is represented by only a large cusp t1 forming the anterolingual portion of the tooth. In seven out of 10 specimens, a small cusp t3 sits on the anterolabial margin; in two others, the cusp is missing and the anterolabial border is marked by only an inconspicuous cingulum; and presence or absence of cusp t 3 in the tenth specimen cannot be determined because the molars are too worn. The complete chevron and chunky posterior portion of the second molar, in size and shape, resembles comparable rows in the first molar. Each third molar has an occlusal pattern that is basically a compacted form of that characterizing the second molar: a conspicuous cusp t1; minute cusp t3 (seen in young rats but unidentifiable in older animals with more worn chewing surfaces); a complete arcuate lamina derived from the total coalesence of cusps t4, t5, and t6; and small posterior chunk either diamondshaped or oblong in outline (formed primarily of cusp t8; cusp t9 is either absent or unidentifiable due to its total incorporation into cusp t8). We could not detect a cusp t7 (on the lingual cingulum between cusps t4 and t8 when present; see Musser and Newcomb, 1983: 333), posterior cingulum (pressed against the posterior margin of cusp t8, if present; see Musser and Newcomb, 1983: 333), accessory cusplets, or longitudinal crests connecting the cusps to form a garland configuration (‘‘stephanodonty’’; see Schaub, 1938, and Misonne, 1969) on any of the upper molars. When present these structures provide complexity to the occlusal planes and in some murines significantly increase chewing surfaces.
Occlusal planes of the lower molars are also comprised of uncomplicated patterns ( fig. 14 View Fig ). An anterolingual cusp coalesced with a smaller anterolabial forms the anteroconid, the most anterior cusp row, which is narrower than the lamina behind it. The second and third are in the shape of narrow chevrons and represent unions of protoconid-metaconid and hypoconid-entoconid, respectively. A roughly triangular posterior cingulum forms the entire posterior fourth of the molar. Except for lacking an anteroconid, occlusal cusp patterns of the second molar resemble patterns seen in the first. The anterior half of each third molar consists of a front chevron formed by union of protoconid and metaconid and behind that a large oblong structure likely representing the complete fusion of hypoconid and entoconid. Anterolabial cusps are absent from second and third molars in most specimens, but we are not certain about others in the sample because their molars are worn to a level such that, if present, these cusps are now indistinct because they have completely coalesced with the adjacent lamina; we did find evidence of a small anterolabial cusp on the third molar of the holotype ( fig. 14 View Fig ). We had the same frustration determining the frequency of posterior labial cusplets on some specimens. This small cone is prominent and forms the anterolabial margins of the posterior laminae of first and second molars in the holotype ( fig. 14 View Fig ). The cusplets are present but inconspicuous on first and second molars in three other specimens (the laminar outline reflects the blending of a cusplet with the anterolabial laminar margin), and are either absent or undetectable in seven rats (because the cusplet has completely coalesced with the adjacent cusp). A posterolabial cusplet clearly does not occur on any third molar in any of the specimens at hand.


| AMCC |
Ambrose Monell Cryo Collection, American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.