
Megalosauridae, Fitzinger, 1843
|
publication ID |
https://doi.org/10.5252/geodiversitas2022v44a12 |
|
publication LSID |
urn:lsid:zoobank.org:pub:231E3A73-1800-4559-A995-013482DEA623 |
|
DOI |
https://doi.org/10.5281/zenodo.6399011 |
|
persistent identifier |
https://treatment.plazi.org/id/03F29C74-FFF4-961E-D6B4-6E93FA9B9CE5 |
|
treatment provided by |
Felipe |
|
scientific name |
Megalosauridae |
| status |
|
Megalosauridae indet.
CRANIAL SKELETON
Premaxilla
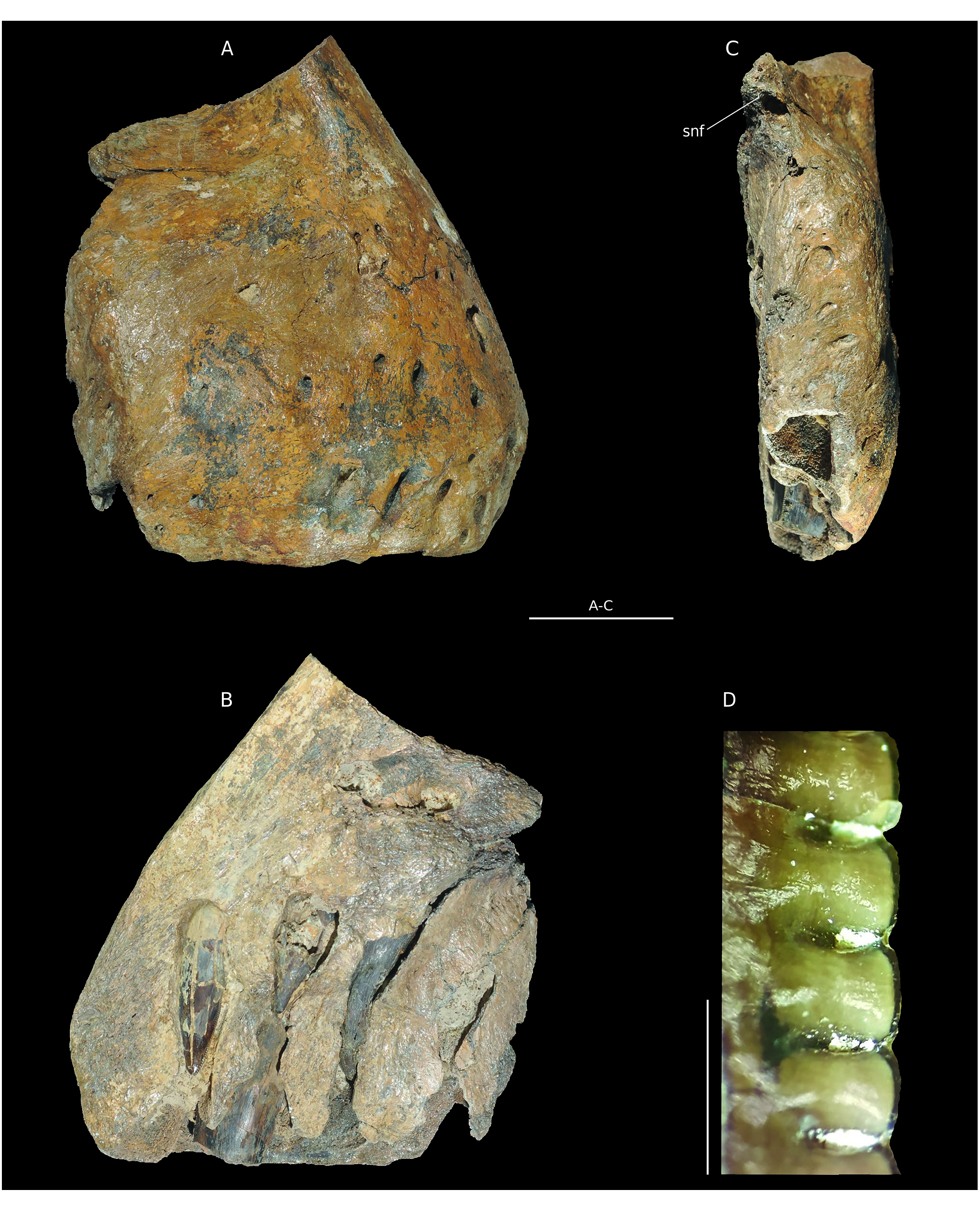
Material. B1, a right premaxilla ( Fig. 2 View FIG ).
Stratigraphy. Oolithe Ferrugineuse de Villers (H15) as defined by Hébert (1860) maintained in Dugué et al. (1998), according to the ferruginous ooliths of the matrix. Lower Oxfordian.
The posterodorsal part of the premaxilla is fractured and the nasal process is missing, clearly cut at its base. The maxillary process is also absent. The medial side is eroded, revealing dental alveoli and replacement teeth ( Fig. 2B View FIG ). The bone is massive. It is 170 mm high under the ventral margin of the nostril and 160 mm long. By comparison with theropods skulls, notably Allosaurus fragilis ( Madsen 1976) , the entire skull length must have exceeded one meter. Similarly to Torvosaurus tanneri Galton & Jensen, 1979 , the premaxillary body is as long as it is wide ( Britt 1991). The lateral side is pierced by more than 15 foramina. Most of them are wider than 5 mm and concentrated in the anteroventral corner of the bone. They are interpreted as perforations for trigeminal nerve branches of the neurovascular system ( Barker et al. 2017). As in Torvosaurus tanneri ( Britt 1991) the anterior part of the bone slopes rearward ( Fig. 2A View FIG ). The medial surface is flat to form the symphysis with the left premaxilla. The width of the bone at this level is 80 mm so that the entire snout width was about 160 mm. A depression linked to the external nares is visible in the posterodorsal corner. Thus, the premaxillary body is quite anteriorly placed relative to the nostril. The angle between the basis of the nasal process and the alveolar margin is about 60°.This is more similar to Eustreptospondylus Walker, 1964 , Dubreuillosaurus valesdunensis ( Allain, 2002) and Torvosaurus than to Sinraptor Currie & Zhao, 1993 or Allosaurus Marsh, 1877 . Posteriorly, the contact facet with the maxilla is visible. It is flat and medially oriented, with three foramina. The subnarial foramen is placed dorsally on the facet ( Fig. 2C View FIG ). On the medial face, the interdental plates are broken so that replacement teeth are visible. However, they do not seem to be fused in their ventral margin. The paradental lamina separates the interdental plates from the body of the premaxilla body. In ventral view, 4 ellipsoidal alveoli are visible. They are mediolaterally oriented, with an erupted tooth in the second one. This tooth is transversally broken at the level of the ventral margin of the premaxilla ( Fig. 2C, D View FIG ). Above this tooth, the apex of a replacement tooth is visible. The first and third alveoli bear only one replacement tooth each. Denticles are mainly visible on the teeth of the first and second alveoli ( Fig. 2D View FIG ). There are seven mesial denticles per five millimetres; they are subrectangular and perpendicular to the crown. However, these dental characteristics are very similar to those of both allosauroids and megalosauroids (except spinosaurids) so that they do not help to differentiate these groups ( Hendrickx et al. 2014).
APPENDICULAR SKELETON
Femora
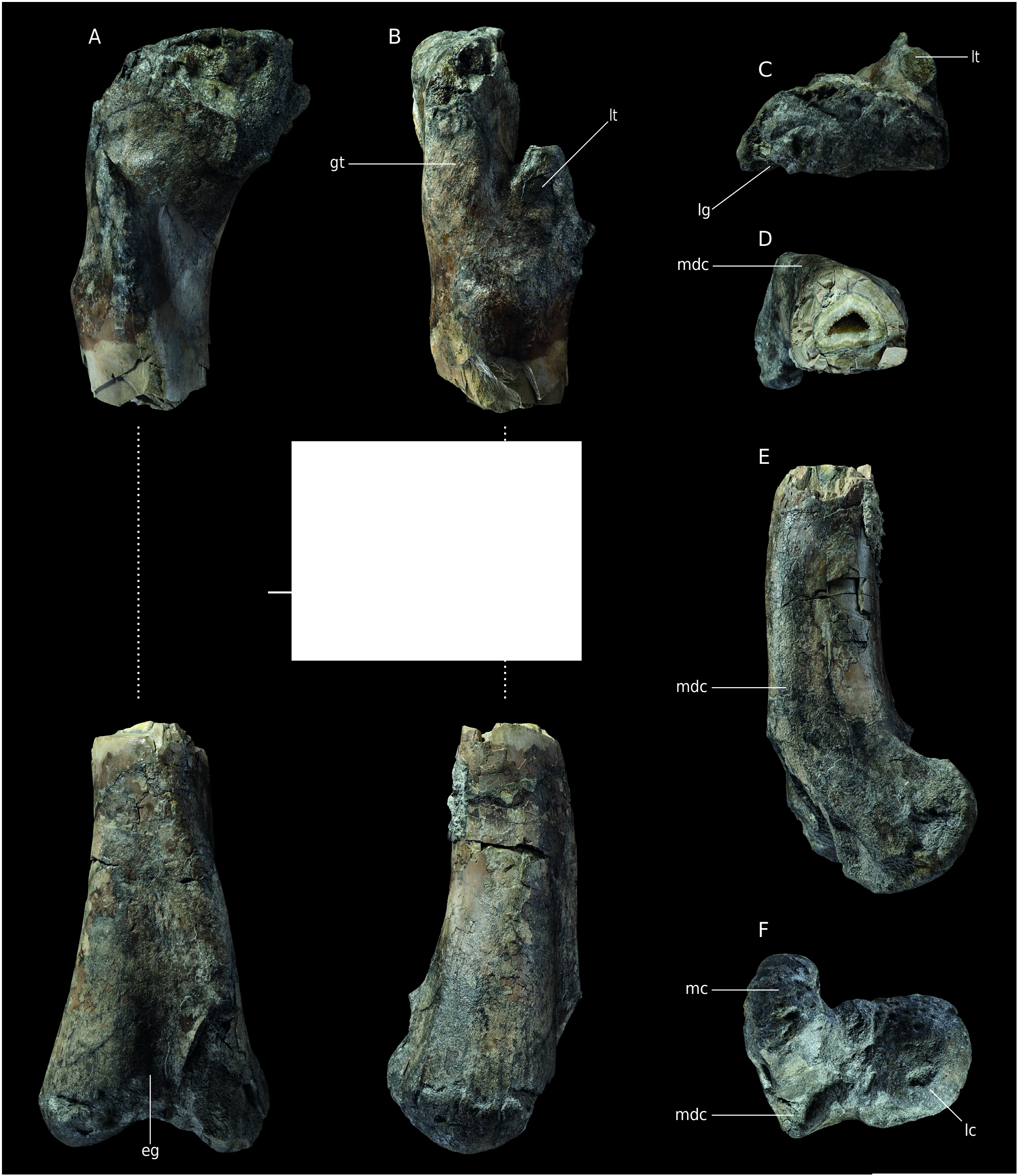
Material. right femur MNHN.F.RJN471( Fig. 3 View FIG ); NMH1, a distal end of a left femur.
Stratigraphy. Found in situ in the Marnes de Villers (H14 level according toHébert [1860] maintained by Dugué et al. [1998]). Lower Oxfordian ( MNHN.F.RJN471). Marnes de Dives Callovian marls according to the Nicolet collection catalogue made by Max Bülow and Michel Rioult (NMH1).
The diaphysis is missing so that the femur MNHN.F.RJN471 is in two parts ( Fig.3A, B View FIG ). Both extremities are quite fractured and eroded, especially the proximal one. Only the base of the articular head is preserved. The proximal part of the greater trochanter is broken, as well as the lesser trochanter.The fourth trochanter is missing because this part of the diaphysis is not preserved. The distal part is broken anteromedially. The crista tibiofibularis is missing, leaving a subrectangular base. The femur is massive and belongs to a very large theropod. The complete bone may have been up to one metre in length.
In posterior view, a ligament groove is visible just lateral to what could be the emplacement of the base of the articular head. Below that structure, in the long axis of the bone a 10 mm long oval foramen is visible. Because the middle part of the diaphysis is missing as well as the head, the orientation of the latter cannot be reconstructed. The greater trochanter is a 55 mm large bony plate on the lateral side of the bone. Its anterior middle margin is slightly convex proximodistally. The lesser trochanter is a thick blade of bone detached from the diaphysis and anteriorly oriented ( Fig. 3C View FIG ). Even though it is damaged, it probably did not reach the proximal margin of the femur, as is the case in basal tetanuran and nontetanuran theropods ( Carrano et al. 2012). A protuberance distally positioned on the trochanter could be a remain of the accessory trochanter, but abrasion makes the observation unclear. A bulge is visible medially to the most distal point of the lesser trochanter, which is interpreted as an attachment for the iliofemoralis muscle ( Hutchinson 2001)
The distal extremity of the femur is 229 mm wide mediolaterally so that it is larger than the other femur (NHM1) described by Buffetaut (1994b). In distal view, both condyles remain visible. The lateral one is rounded and the medial one is narrower and elongate ( Fig. 3F View FIG ). The medial condyle bears deep and wide striations (one posterior and one anterior) but they may have been exaggerated by erosion. The medial condyle is anteroposteriorly oriented, but the articular facet seems more posterior. As a result, in medial view, the bone takes the shape of a reversed J ( Fig. 3E View FIG ). The medial condyle bears a striking feature above the broken area: a very thick and high mesiodistal crest extending on the midline of the distal diaphysis. The crest peaks at about 40 mm relative to the lateral side. This structure makes the cross section of the diaphysis triangular ( Fig. 3D View FIG ). This is very different from the condition in Streptospondylus, in which this area is flat. There is no depression for the femorotibialis externus ( Hutchinson 2001; Carrano et al. 2012) on that crest in contrast to what is seen in allosauroids ( Benson 2010; Carrano et al. 2012; Hendrickx & Mateus 2014). Moreover, the crest is rounded and thick in contrast with some allosauroids such as Allosaurus and Sinraptor ( Currie & Zhao 1994) . A similar morphology of the structure is mentioned by Hendrickx & Mateus (2014) on a distal fragment of a femur referred to Torvosaurus . This hypertrophied mesiodistal crest is present in the two species of the genus, but it is reduced as compared with MNHN.F.RJN471 ( Hendrickx & Mateus 2014) insofar as it does not make the cross section triangular ( Siegwarth et al. 1997). In Megalosaurus Buckland, 1822 the mesiodistal crest is poorly developed ( Benson 2010; Hendrickx & Mateus 2014). The extensor groove is only visible in its most proximal part. The flexor groove is convex and 30 mm wide.
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Tetanurae |
|
Family |