
Jania rosea (Lamarck) Decaisne
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.606.2.1 |
|
DOI |
https://doi.org/10.5281/zenodo.8226471 |
|
persistent identifier |
https://treatment.plazi.org/id/03F187C8-FF9D-FFD5-FF4C-FA388B90FE15 |
|
treatment provided by |
Plazi |
|
scientific name |
Jania rosea (Lamarck) Decaisne |
| status |
|
Jania rosea (Lamarck) Decaisne
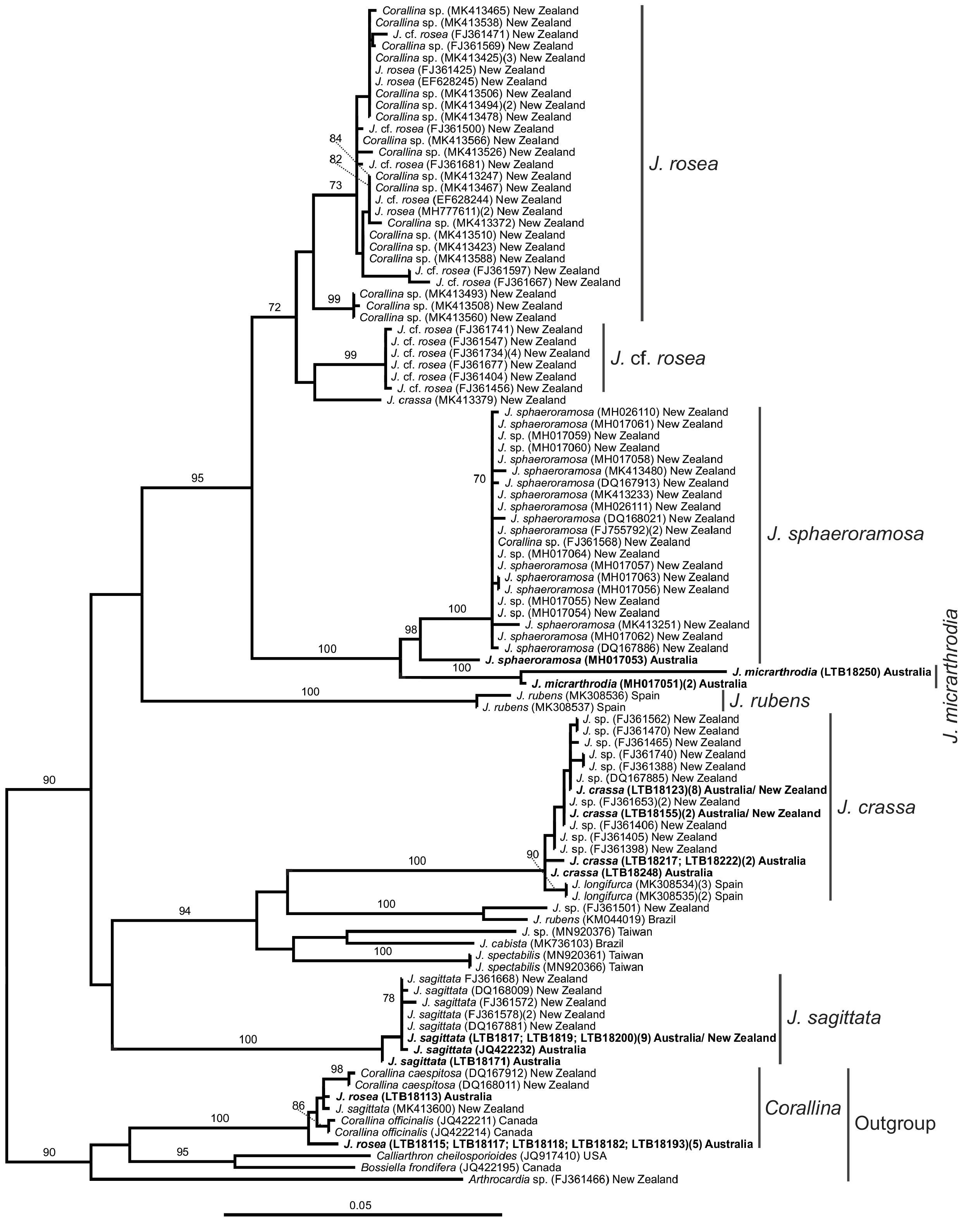
— Jania rosea is extensively recorded across temperate Australia (Australian Virtual Herbarium 2022) and other parts of the world (see AlgaeBase; Guiry and Guiry 2022) such as California ( Miller 2012), Brazil ( Buys and Gurgel 1998), Indonesia ( Atmadja and Prud’homme van Reine 2010), and Sub-Antarctic Islands ( Papenfuss 1964). Johansen and Womersley (1986) reported a wide range of variation in thallus morphology in J. rosea . Specimens can grow as purely dichotomously divided thalli, resembling several other Jania species and varieties such as J. pedunculata var. pedunculata ( Harvey et al. 2020, fig. 8), J. pedunculata var. adhaerens ( Harvey et al. 2020, fig. 17), J. micrarthrodia ( Harvey et al. 2020, fig. 6), and J. capillacea Harvey ( Mendoza-González et al. 2014, figs. 30-32). Jania rosea can also display a defined slightly compressed main axis and pinnate branches, usually when growing attached to rocks in more exposed sites ( Farr et al. 2009, Harvey et al. 2020), or forming well-defined main axis with opposite (or even radially and whorled) branching, resembling some delicate Corallina species (e.g., Calderon et al. 2021).
Australian J. rosea morphotypes have not been recognized as distinct species due to excessive phenotypic plasticity and the high level of observed overlap between morphotypes ( Johansen and Womersley 1986, Harvey et al. 2020). Farr et al. (2009) working with New Zealand specimens, found evidence to differentiate two closely related J. rosea clades also on the basis of morphological evidence (i.e., feather versus bottlebrush morphotypes). However, Farr et al. (2009) treated these two distinct genetic entities as a single species, and emphasized ‘that further work is required, and additional species are likely to be recognized in the future’.
Cox 1 phylogenies and SDM results recognized that J.rosea from Australia is a different species and phylogenetically distantly related to the two species passing under the name J. rosea in South Africa ( Kogame et al. 2017). Cox 1 molecular data showed that the lineage named J. rosea from South Africa was resolved as a paraphyletic group closely related to J. subulata (Ellis and Solander) Sonder from Hawaii ( Fig. 3 View FIGURE 3 ). In this study the psb A phylogenetic results showed that one J. rosea specimen (LTB18113) formed a clade with C. caespitosa from New Zealand and another five specimens formed a distinct clade with C. berteroi from Australia (LTB18115, LTB18117, LTB18118, LTB18182, and LTB18193), Chile (MZ262616, MZ262579) and California, USA (MZ262623). These data evidence how challenging the morphology-based taxonomy can be and that some Corallina and Jania morpho-anatomical characters overlap.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |