
Staphylinus
|
publication ID |
https://doi.org/10.5281/zenodo.283045 |
|
publication LSID |
lsid:zoobank.org:pub:2F9DF810-4E89-4BE9-BBB2-EC11D64AF272 |
|
DOI |
https://doi.org/10.5281/zenodo.5690233 |
|
persistent identifier |
https://treatment.plazi.org/id/03F187C7-2435-FFAC-FF7A-F92CFB7CF9D4 |
|
treatment provided by |
Plazi |
|
scientific name |
Staphylinus |
| status |
|
Generic diagnosis of mature larvae. The larval description of S. erythropterus presented in this paper is not in agreement with the earlier one by Boháč (1982). Therefore, these literature data were not applied in further diagnostic analysis. The morphological data regarding S. erythropterus presented below combined with the data extracted from the superficial description of S. cesareus ( Pototskaya 1967) were used to determine the diagnostic characters of genus Staphylinus . They are as fallow: frayed setae absent; anterior margin of labrum with 9 very weekly visible, slightly separated to each others, low teeth; mandible with four setae; small tooth on mandible; stalk of apotome wide; ligula stump-shaped with two pairs of setae; hypopharynx membranous and thickly pubescent; stipes of maxilla with hair-like cuticular processes; mala short (2.5 times shorter to segment I of maxillary palp); maxillary and labial palp three- and two-segmented, respectively; foretibia with comb of bifid setae; tarsungulus bisetose; cervicosternum triangular divided at least partially; urogomphi longer to pygopod.
Generic diagnosis of pupa. The description of pupae of S. erythropterus given by Szujecki (1960) is incomplete. The author described and illustrated the pupae exclusively based on one specimen collected in the field. Due to this, some significant morphological structures could be broken, and therefore not recorded or illustrated. The combination of characters permitting the distinguishing of pupae of Staphylinus from other known pupae of subtribe Staphylinina ( Creophilus , Ocypus , Ontholestes , Platydracus , Tasgius ) is as follows: the length of hind leg reaching half length of the 5th (morphologically 7th) well visible from ventral side abdominal segment; the presence of pair setae on three terminal, abdominal segments (VII – IX); the number of setiform projections on pronotum (19 – 25); antenna, not protruding half length of the shortened elytra; the body length: 9 – 10.2 mm ( Dajoz & Caussanel 1968; Schmidt 1994; Staniec 2004; Staniec & Pietrykowska 2005; Staniec et al. 2009). A brief description and sketchy figure of pupal second member of Staphylinus—S. cesareus were given by Boháč (1982). Considering a scant, intrageneric comparison, S. erythropterus differs from S. cesareus in the tarsi reaching half length of the 5th well visible abdominal segment ( S. cesareus have tarsi protruding to anterior margin of the 3rd abdominal segment).
Description of mature larva and pupa of S. erythropterus (adult habitus and aedeagus as in Figs 62, 65, 66)
Larva (L2) (larval habitus as in Figs 4–6 View FIGURES 1 – 11 ) (only selected features)
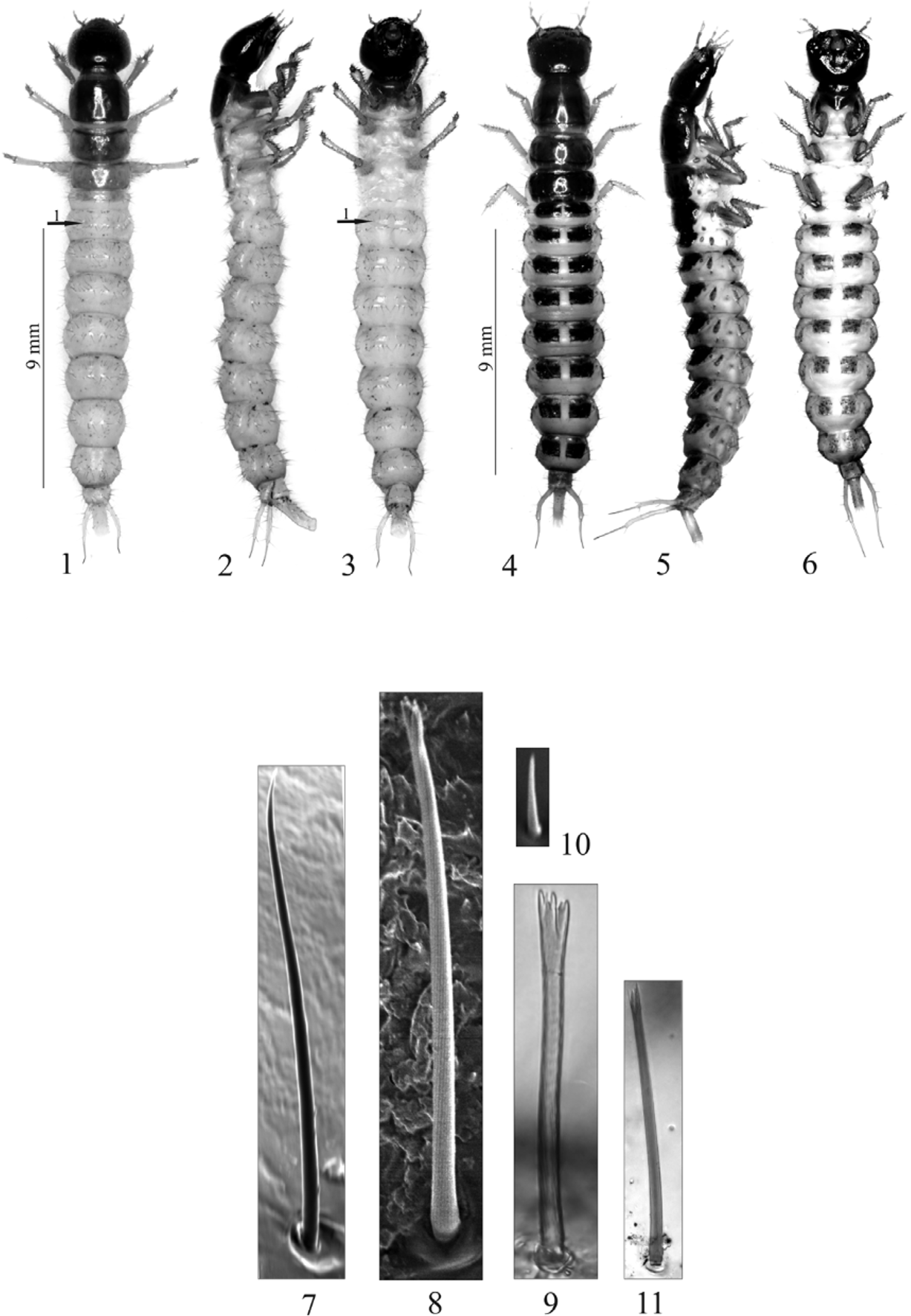
Body length: 16.9 – 21.5 mm (mean 18.4 mm); head width (HW1): 2.0 – 2.7 mm (mean 2.4 mm), head width (HW2): 2.3 – 2.9 mm (mean 2.5 mm); head length: 1.6 – 2.9 mm (mean 2.1 mm); pronotum width: 2.0 – 2.7 mm (mean 2.3 mm). Colour: head dark brown, mandibles almost black, antennae, maxillae, labium and legs yellowish brow, pronotum and mesonotum brown, metanotum dark brown, abdominal tergites I – VIII almost black, tergite IX and sternites I – IX grey, urogomphi and apical part of segment X dark yellow, all setae (macro and micro) brown and simple. Body elongated, cylindrical first abdominal segments narrower than second one, segments II – V more or less equal in length then gradually narrowed to the terminal segment of the body.
Head (Figs 14, 15, 17, 18, 20, 22, 26, 27, 30, 31, 35–37, 39, 42): lateral margins almost parallel, widest about at the stemmata level (Fig. 15). Each side of head with four stemmata in a cluster, three stemmata almost of equal size, the hind one distinctly bigger than others (Fig. 14). Epicranial part (E) with 20 ( 2x10) macro setae located symmetrically in relation to epicranial suture (Fig. 15). Nasale (Na) without asymmetrically setae (Fig. 18); anterior margin of nasale with 9 weakly visible, slightly separated each other, low and wide teeth; all teeth (Mt, Pmt, Lt1 – 3) equal in length (Fig. 17). Ventral side of head with 14 ( 2x 7) macro setae located symmetrically in relation to ventral suture, without micro setae (Fig. 20). Apotome (Ap) (Figs 20, 22) in broad outline triangular, with wide stalk, slightly extending beyond tentorial pits.
Antenna (Figs 26, 27) 4-segmented, length ratio of segments I – IV 1.2:2.5:1.7:1.0, respectively; segment I as wide at the base as long; segment II 4.2 times as long as wide, with two pores (one dorsally, one ventrally); segment III 3.5 times as long as wide in the widest place, with three macro setae laterally, three sensory appendages (Sa) – one biggest acorn-shaped and two tiny (Fig. 26); segment IV about 3.5 times as long as wide, about 3.9 times as long as acorn-shaped sensory appendage of segment III, with 3 setae short or long (Figs 26, 27) and 4 solenidia (So) apically (Fig. 26). Mandible (Figs 30, 30A, 31) slender and narrow relatively, with 4 setae (coded: 1 – 4) at the outer margin, among them: one (coded: 3) distinctly longer than others and one tiny (coded: 1 or 2) very often invisible, 1 pore dorsally; inner margin with two teeth–tiny, poorly visible in basal part (Fig. 30A) and bigger one, obtuse, low in about 1/3 length from the base. Maxilla (Figs 35 – 37): length ratio of cardo (Cd) and stipes (Stp) 1:2.5; cardo 1.3 times as wide as long bearing 1 seta ventro-laterally; stipes 2.6 times as long as wide with 9 setae (4 near outer margin, 3 near inner margin, and 2 near mala) and region of hair-like cuticular processes laterobasally (Fig. 36A). Mala (Ma) (Figs 35, 35A) finger-shaped, slightly narrowed apically, with 2 setae, 3 micro sensory appendage apically and 3 pores; length ratio of mala and segment I of maxillary palp: 1:2.4. Palpifer (Pf) (Fig. 35) with 1 pore and 1 seta ventrally. Maxillary palp (Pm) 3-segmented, slim relatively, segments I and II almost equal of width; length ratio of segments I–III: 1:1.1:1.2, respectively; segment I 3.7 times as long as wide, slightly and gradually winded to its apical, with 2 pores ventrally; segment II 4.9 times as long as wide, slightly winded to its apical, with 2 setae (1 at the outer margin near apical, 1 at the inner margin in 1/3 length from the base) and 2 pore near apical (Fig. 36); segment III slightly narrower than others, 8.8 times as long as wide, with 1 digitiform sensory appendage basally on outer margin, 2 pores near apex and a few micro sensory appendages apically, length ratio segment and digitiform sensory appendage: 7.5:1 (Fig. 37). Hypopharynx: dorsal side of labium membranous and thickly pubescent, apex sinuate (Figs 39, 39A). Prementum (Fig. 42) consists of two parts: anterior–wider and membranous with ligula (Lg) and a pair of labial palps, and posterior–narrower, sclerotized, narrowed basally with 4 setae (2 long anteriorly, 2 short laterally); ligula short and rounded, with 4 micro setae, separated from prementum by wide sclerotized stripe (Fig. 42A). Labial palp (Lp) 2-segmented (Fig. 42); length ratio of segments I – II 1.2:1, respectively; segment I slightly and gradually widened to the apical, 4.6 x as long as wide in the widest place, with 1 pore at the inner margin; segment II slightly narrower than the first one, gradually narrowed to the apical, 5.4 x as long as wide in the widest place, with one pore near apical and few micro sensory appendages apically.
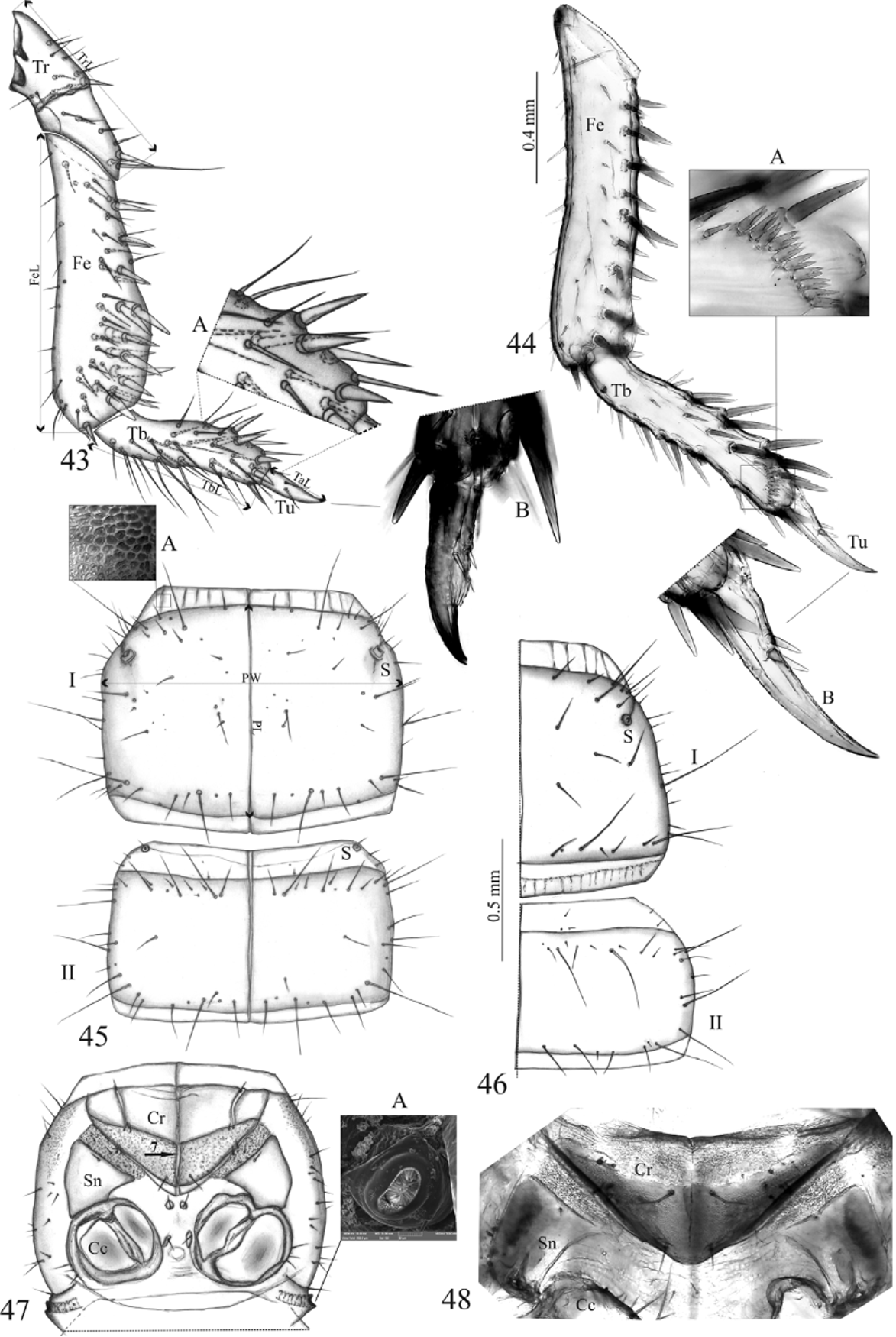
Thorax ( Figs 44, 46, 48 View FIGURES 43 – 48 ). Foreleg ( Fig. 44 View FIGURES 43 – 48 ): femur (Fe) with about 45 setae (20 thick, spine-shaped) and 2 – 3 pores; tibia (Tb) with 30 thick, spine-shaped setae (3 short) and comb of 10 – 13 bifid setae near apex ( Figs 44, 44 View FIGURES 43 – 48 A); tarsungulus (Tu) with 2 spine-shaped setae ( Fig. 44 View FIGURES 43 – 48 B); length ratio of profemur, protibia and protarsungulus 3.3:2.2:1, respectively. Pro-, meso, and metanotum with mid-longitudinal ecdysial line; tergite I with a pair of sensilla (S) antero-laterally; chaetotaxy of pro- and mesonotum as in Fig. 46 View FIGURES 43 – 48 ; chaetotaxy of meso- and metanotum identical. Cervicosternum (Cr) triangular, divided not completly by ecdysial longitudinal line ( Fig. 48 View FIGURES 43 – 48 ).
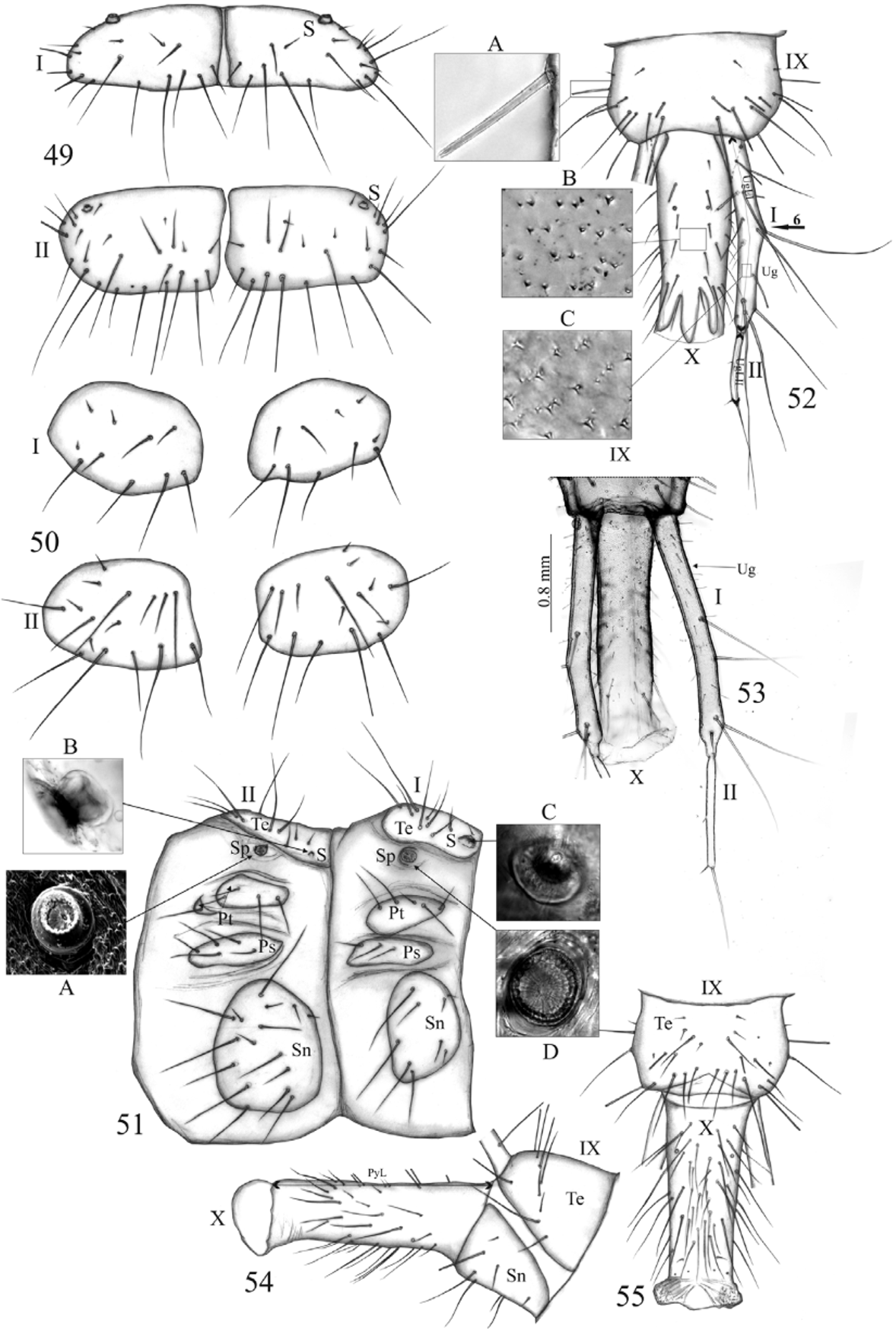
Abdomen ( Fig. 53 View FIGURES 49 – 55 ). Urogomphi (Ug), each one with two clearly visible segments ( Fig. 53 View FIGURES 49 – 55 ): segment I slightly arched to inner side with about 30 setae (about 25 short, 5 long) and a few pores; segment II straight with 3 setae (two micro, one long apically); length ratio of segments I, II of and apical seta 3.0:1.3:1.0, respectively. Urogomphi (without seta apically) distinctly longer than pygopod; length ratio of segment I, II of urogomphi and pygopod 2.3:1:2.3.
Pupa (habitus as in Figs 56–60 View FIGURES 56 – 60 ) [supplement to description of Szujecki (1960)]. Body length: 9.04 – 10.16 mm (mean 9.59 mm); body width: 4.31 – 4.80 mm (mean 4.60 mm); head width: 2.94 – 3.36 mm (mean 3.17 mm); head length: 3.13 – 3.45 mm (mean 3.29 mm); pronotum width: 2.79 – 3.11 mm (mean 2.98 mm), pronotum length: 2.89 – 3.16 mm (mean 3.01 mm); colour bricky-brown, body stocky.
Head almost as long as wide ( Figs 56, 57 View FIGURES 56 – 60 ), antennae reaching about one four of the elytra length, labrum about 1.6 times as broad (at the base) as long; mandible crossed on their apexes. Anterior margin of pronotum with 19 – 25 projections (NPP) (13/12, 12/13, 12/12, 13/11, 11/12, 10/12, 10/11, 10/10, 9/11, 8/12, 10/9 on sides in individuals) ( Fig. 57 View FIGURES 56 – 60 B). Wings reaching the posterior margin of 1st (morphologically 3rd) clearly visible from ventral side, abdominal segment ( Figs 56, 57 View FIGURES 56 – 60 ). Each middle tibiae with at least 15 outlines of protuberances ( Fig. 57 View FIGURES 56 – 60 A), hind tibiae with 4–5 clearly visible protuberances on tarsi. Tarsi of hind legs reaching half length of the 5th (morphologically 7th) well visible from ventral side abdominal segment ( Fig. 56 View FIGURES 56 – 60 ). Abdominal tergite I 1.6 times longer than tergite II. Tergites I – IV with tuberculate, functional spiracles ( Fig. 58 View FIGURES 56 – 60 ), the first pair situated more laterally than the rest, moderately protruding; tergites V – VIII with externally visible, but apparently atrophied spiracles ( Fig. 58 View FIGURES 56 – 60 A). Segments VII – VIII each bearing a pair of setiform projections on sides ( Fig. 58 View FIGURES 56 – 60 B). Setiform projections on segment VII longer to VIII. Terminal sternite with well marked sexual dimorphism: female pupa with double gonotheca, male pupa with single gonotheca ( Figs 59, 60 View FIGURES 56 – 60 ), each sex with a pair of curved, terminal prolongation with numerous, cuticular processes ( Fig. 60 View FIGURES 56 – 60 A).
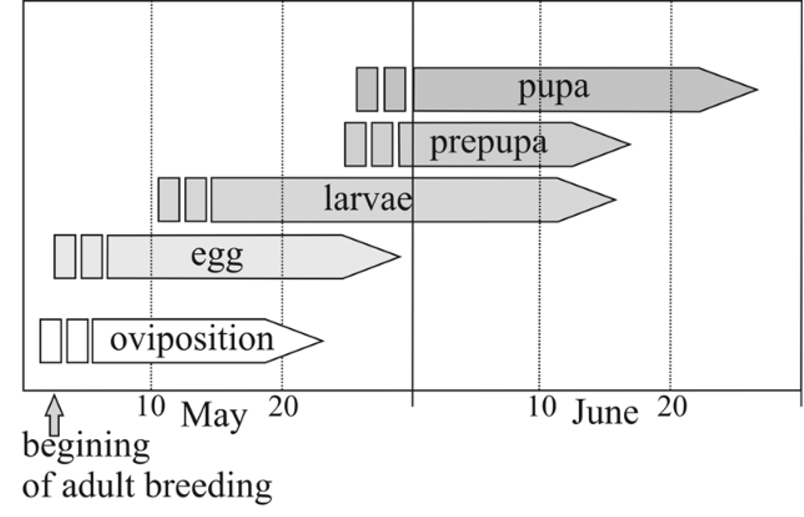
Notes on the life history of S. erythropterus in laboratory condition. During the rearing, conducted from 3 May to 15 June 2011, the insects were able to fully complete their life cycle. Immediately after catching, imagines started copulation that repeated with intervals for a period of 6–12 days (from 5 to 17 May). The female oviposition lasted from 6 to 21 May (15 days in total). The time of the copulation stage coincided with the oviposition in laboratory conditions. This suggests that the actual time of oviposition (in field conditions) could have started before 3 May. During the oviposition period, insects hid in the ground and laid eggs every day. One female laid 1 – 2, sporadically 3 eggs per day. These data, however, may not be reliable, because they result from rearing of only two females (the third female died after laying 3 eggs). Eggs were laid separately, distributed in the soil filling the rearing containers. During the oviposition (15 days) one female laid a total of 26 eggs.
The period of embryonic development varied from 8 to 10 days (mean 9 days). In the laboratory conditions the larval development lasted from the middle of May (14.05) to the middle of June (12.06). The development of particular larval stages ranged from 5 to 7 days (mean 6 days) for the first stage (L1), and 7–9 days (mean 8 days) for the second (L2). The time of occurrence of both larval stages lasted 4 weeks. Shortly before pupation, S. erythropterus larvae, as usually most of the known larvae of Staphylinini , were becoming less and less active. The mean duration of development of the prepupae and pupae was 2 and 12 days, respectively. Prepupae and pupae were observed in the laboratory from the end of May (29.05) to June (26.06). After the metamorphosis, the mature specimens of the new generation came out onto the soil surface in the first decade of June (10.06). Their appearance coincided with the presence of previous-generation specimens.
The complete duration of the development of S. erythropterus under laboratory conditions, from the egg until the emergence of the adult form of a new generation, was 36 days on the average ( Fig. 67 View FIGURE 67 ).
Remarks on the inherency of two larval instars of S. erythropterus unique within Staphylinini . The present paper provides a new insights into the life cycle of representatives of tribe Staphylinini . S. erythropterus has only two larval instars, in contrast to three instars observed in the case of other closely related species: Acylophorus wagenschieberi , Creophilus maxillosus , Ocypus fulvipennis , Philonthus rubripennis , Platydracus latebricola , Quedius brevicornis ( Dajoz & Caussanel 1968; Staniec 2003; Staniec 2005; Staniec & Pietrykowska- Tudruj 2007; Staniec et al. 2009). The following evidences lack of the third larval instar in the life cycle of S. erythropterus : differences in the size of morphological structures between successive larval stages, short time of the total duration of the life cycle, amounting to 13,6 days, and the presence of only two larval exzuviae (belonging to L1 and L2) in the rearing chambers. Table 3 presents the comparison of the size of the most important morphological structures: head width and length, as well as pronotum width and body length of the first and second larval stage and prepupa. The convergence of the size of all the selected structures in the second larval instar and prepupa strongly suggests that this (second) larval stage is the last one. The distinction of two larval stages in the life cycle of S. erythropterus is in accordance with the determinations by Tikhomirova & Melnikov (1970), and contradicts those by Boháč (1982). Based on laboratory rearing, Boháč distinguished and described three larval stages. The comparison of the head width of all larval stages, presented herein, however, firmly denies his statement, and confirms the lack of the third larval stage.
It is interesting, how the process of embryonisation is reflected on the larval morphology. In their study on the late embryogenesis of Staphylinus, Tikhomirova and Melnikov (1970) maintain that the first larval stages are the “missing” ones. According to the authors, the process of embryonisation has gone so far that the first larval stage has been completely included in the egg. Newly hatched larvae of Staphylinus have well sclerotised slerites and developed chaetotaxy. Their morphology is similar to that of the second larval instars of other known larvae of Staphylinini . Our evaluation of larval characters, however, is not in complete accordance with the the statement of Tikhomirova and Melnikov (1970). In the case of Staphylinini , as far as their immature stages are known, the newly hatched larvae of S. erythropterus have characters also observed in both the second and first larval stage ( Staniec 2003; Staniec et al., 2009). The main features of newly hatched larvae making them morphologicaly similar to the first stage are the following: lack of comb of bifid setae on the tarsungulus; a lower number of setae (in comparison to the last larval instar) on the femur, tibia (27 and 22, respectively) and segment I of the urogomphi (about 18); the presence of rod-shaped setae on the abdominal tergites. Compared with the second larval stage, newly hatched larvae are similar in terms of the body size. They also have the same number of sensory appendages and solenidia on the third antennal segment (3 and 1, respectively) as well as setae on tarsungulus (two setae).
In conclusion, the newly hatched larvae of S. erythropterus combine morphological features, observed in the first and second larval instars of other known larvae of Staphylinini . The tendency of embryonisation of S. erythropterus may be related to the intensification of predation of this large and very active staphylinid ( Tikhomirova & Melnikov 1970).
Concluding remarks. Platydracus Thomson, 1858 was originally described as a separated genus. Until 1956, however, almost all authors considered the taxon a subgenus of the genus Staphylinus Linné. Even though Coiffait (1956) re-elevated Platydracus to the genus level, some authors still consider Platydracus a subgenus. The taxonomic history of both of the genera shows they are closely related. As usually, all these taxonomic transfers were done besed exclusively on adult feateures. Our study enriches the current database of adult characters with new, larval ones. Numerous significant characters suggesting the genus level of Platydracus , and the distinction between the larvae of P. latebricola and S. erythropterus presented in table 2, seem definitely confirm the validity position of the categorization of the species as separate genera.
FIGURES 12–22. Mature larva of P. latebricola (12, 13, 16, 19, 21) and S. erythropterus (14, 15, 17, 18, 20, 22). (12, 15, 19, 20) Head, in dorsal aspect (12, 15*) with microstructure (A), ventral aspect (19, 20*); (13, 14) stemmata; (16, 18) nasale; (17) anterior part of nasale; (21, 22) apotome and tentorial pits. Abbreviations: Ap—apotome, E—epicranial part, Gp—glandular pit, Lt—lateral teeth, Mt—median tooth, Na—nasale, P—posterior part, Pmt—paramedian tooth, Tp—tentorial pit, *—right or left half.
FIGURES 23–31. Mature larva of P. latebricola (23–25, 28, 29) and S. erythropterus (26, 27, 30, 31). (23, 26) Apical part of right antenna in dorsal aspect; (24, 27) right antenna in dorsal aspect with version of apical part of segment IV bearing long setae (27A); (25) region of sensory appendages of antennal segment III in ventral aspect; (28) epipharynx; (29, 30) right mandibles with regions of two short sutures convergent to furrow (29A) in dorsal aspect; (31) basal part of mandible. Abbreviations: I–IV—antennal segments, Sa—sensory appendage, So—solenidia.
FIGURES 32–42. Mature larva of P. latebricola (32–34, 38, 40, 41) and S. erythropterus (35–37, 39, 42). (32, 35) Region of mala in ventral aspect; (34, 36) right maxilla in dorsal aspect with inner margin of stipes (34A, 36A); (33, 37) segment III of maxillary palp; (38, 39) hypopharynx with anterior part (39A); (40, 42) labium with ligula (42A) in ventral aspect; (41) segment II of labial palp in dorsal aspect. Abbreviations: I–III—maxillary palps segments, I–II—labial palps segments, Cd—cardo; Lg—ligula, Lp—labial palp, Ma—mala, Mp—maxillary palp, Pf—palpifer, Pmnt—prementum, St—stipes.
FIGURES 61–66. Adult of P. latebricola (61, 63, 64) and S. erythropterus (62, 65, 66). (61, 62) habitus; (63–66) aedeagus in ventral aspect (63, 65) and in lateral aspect (64, 66).
TABLE 2. Principal morphological differences between larvae of Platydracus latebricola (L3) and Staphylinus erythropterus (L2).
Antennal segment II, shape stout (twice as long as wide) slim (four times as long as wide) (Fig. 24) (Fig. 27) Mandible, furrow along inner margin present (Fig. 29) absent Maxilla, length ratio of cardo and stipes 1:1.8 (Fig. 34) 1:2.5 (Fig. 36) Three sensory appendage on the apex of mala, different equal length (Fig. 32) (Fig. 35A) Segments of maxillary palp, length second segment longer the others all segments almost of equal length
(Fig. 35) (Fig. 36) Position of two setae on segment II of the same level (Fig. 34) different level (Fig. 36) maxillary palp
Sides of hypopharynx sclerotized (Fig. 38) membranous and pubescent (Fig. 39) Segment I of labial palp and II of maxillary stout, widened apically (Fig. 40) slim, sides parallel (Fig. 42) palp, shape
Foreleg, length ratio of femur, tibia and 4.7:2.8:1 ( Fig. 43 View FIGURES 43 – 48 ) 3.3:2.2:1 ( Fig. 44 View FIGURES 43 – 48 ) tarsungulus
Foretibia, simple setae (except spine like) present ( Fig. 43, 43 View FIGURES 43 – 48 A) absent
Developmental stage Head width Head length Body length Pronotum width (HW2) (HL) (BL) (PW)
First larval instar rate 1.78–2.09 1.34–1.66 11.8–13.5 1.37–1.80 mean 1.97 1.53 12.4 1.58
Second larval instar rate 2.29–2.89 1.91–2.12 15.7–21.5 2.19–2.71 mean 2.48 2.04 18.5 2.35
Prepupa rate 2.36–2.53 1.95–2.38 16.9–19.7 2.19–2.58 mean 2.46 2.13 18.1 2.34

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Staphylinini |
|
SubTribe |
Staphylinina |
|
Genus |