
Paramonohystera sinica, Yu & Xu, 2014
|
publication ID |
https://doi.org/ 10.1080/00222933.2014.953224 |
|
DOI |
https://doi.org/10.5281/zenodo.4328091 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE3950-5F7E-E13E-85BB-FBAEA3BCFDCA |
|
treatment provided by |
Carolina |
|
scientific name |
Paramonohystera sinica |
| status |
sp. nov. |
Paramonohystera sinica View in CoL sp. nov.
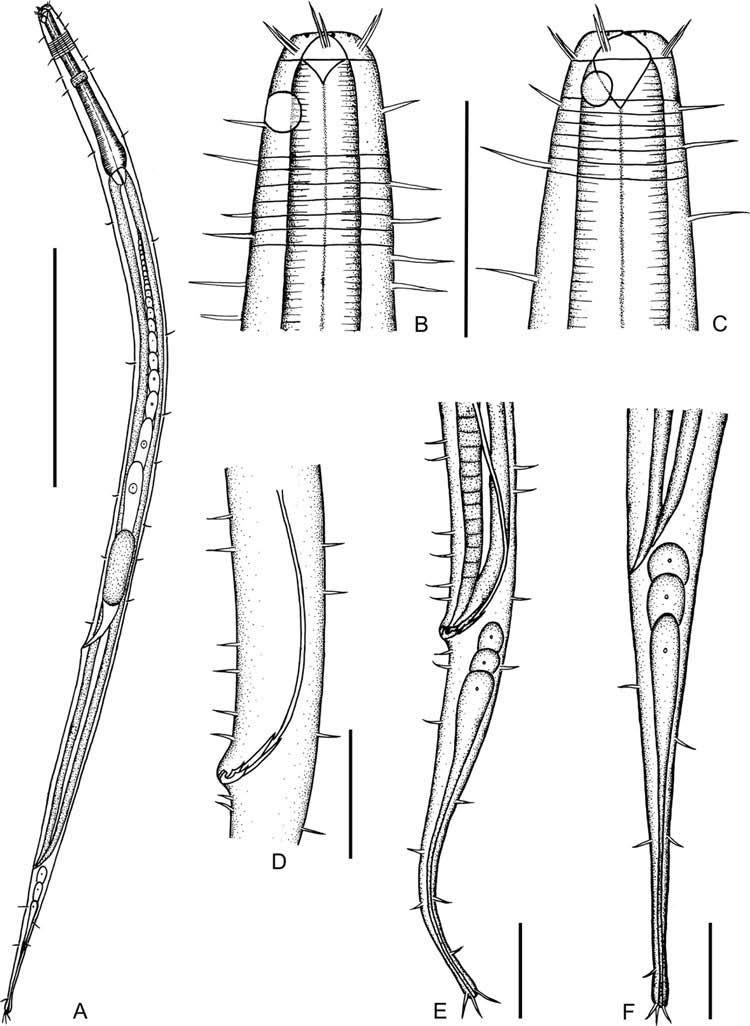
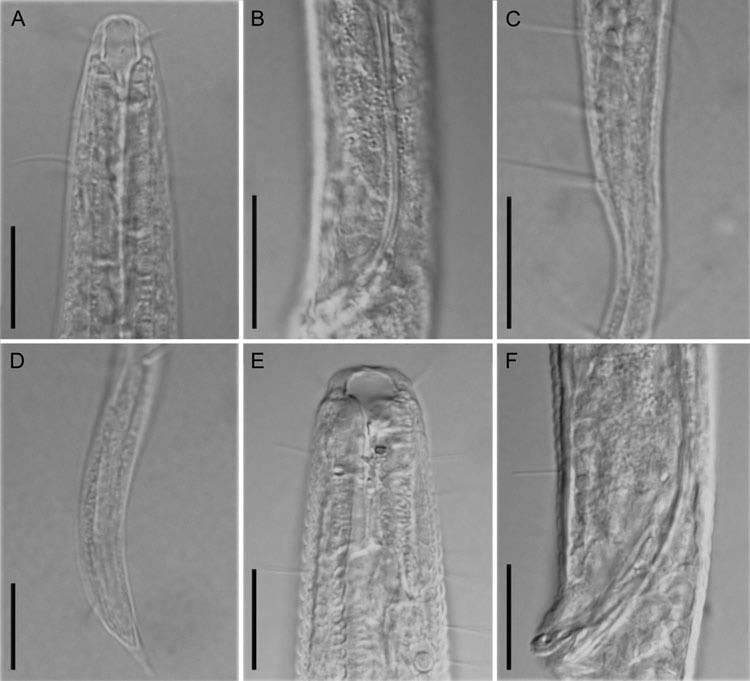
( Figures 2A–F View Figure 2 , 3E, F View Figure 3 ; Tables 2–4)
Diagnosis
Body length about 933–1127 μm. Twelve cephalic setae, smooth and 7–9 μm long. Amphids circular, 6–7 μm across. Cervical setae numerous and up to 14 μm long. Spicules arcuate and slender, about 79–88 μm long and 4.0–4.4 a.b.d. Tail conicocylindrical, 114–146 μm long and 5.7–6.6 a.b.d.
Type material
Five males and four females were measured and studied.
Holotype. One male on slide DH 2-7-02.
Paratypes. Three males and three females on slide DH2-7-02; one male and one female on slide DH2-7-01.
Type locality and habitat
Muddy sediment at Station DH 2-7 (31°01' N, 126°00' E) in the East China Sea; water depth about 70.6 m, water temperature at the sediment–water interface 13.4°C, salinity 33.6, median particle diameter 8 μm, silt-clay 93.6%, organic matter 1.0 GoogleMaps %.
Etymology
The New Latin adjective sinicus (of China) refers to the country where the species was discovered.
Description
Males. Body cylindrical and gradually tapering towards tail end, with head region narrower than body trunk; 933–1023 μm long and 29–34 μm wide at maximum body diameter. Head 13–16 μm wide. Cuticle with coarse annulations visible throughout body, about 2–3 μm at intervals. Many somatic setae scattered all over body, slightly denser and up to 14 μm long at cervical region. Buccal cavity large with hemispherical cheilostome and conical pharyngostome, 7–10 μm wide and 6–11 μm long, both in an average of 8 μm. Anterior sensilla, arranged in two circles: the anterior one composed of six labial papillae, usually difficult to observe; the posterior one with 12 cephalic setae in six pairs, each pair composed of a shorter and a longer seta, the shorter setae 5– 7 μm long, and the longer ones about 7–9 μm long. Amphidial fovea round, 6–7 μm in diameter and 33–46% c.b.d., anterior border of fovea 6–10 μm from anterior body end ( Figure 2B View Figure 2 ). Pharynx cylindrical, slightly widened at base, 153–159 μm long (about 16% of total body length). Pharyngo-intestinal junction with small triangular cardia. Nerve ring located near the middle of pharynx, 75–89 μm distant to anterior body end. Excretory pore and ventral gland not observed. Tail conico-cylindrical, 114–130 μm long and 5.7–6.6 a.b.d., with cylindrical part occupying about one-third of tail length. Three terminal setae, about 7 μm long. Three caudal glands in tail region ( Figure 2E View Figure 2 ).
P. levicula ( Lorenzen 1973) Theristus (Daptonema) leviculus Valid ; spicules 3.4 a.b.d. ( Lorenzen 1972; Gerlach and Lorenzen in Gerlach and Riemann 1973; Lorenzen Riemann, 1973; Theristus 1974 , 1977) ( Daptonema ) levis Lorenzen, 1972
P. longicaudata Timm, 1963 Paramonhystera longicaudata Likely a member of Timm (1963) Timm, 1963 Daptonema ; spicules only 1.5 a.b.d.
P. megacephala Steiner, 1916 Monhystera (Paramonohystera) Valid ; type species, spicules 6.0 ( Filipjev 1918; Wieser 1956; megacephala Steiner, 1916 ; a.b.d. Gerlach and Riemann 1973) Paramonhystera megacephala Steiner, 1916 ; Paramonhystera (P.) megacephala Steiner, 1916
P. micramphis Schuurmans Stekhoven , – Species inquirenda; known ( Wieser and Hopper 1967;
1950 from females and juvenile Chen and Vincx 2000; only Venekey et al. 2014)
P. mutila Lorenzen, 1973 Stylotheristus mutilus Now a member of ( Lorenzen 1973, 1977) ( Lorenzen 1973) Lorenzen, Stylotheristus ; spicules <1 a. 1977 b.d.
P. mystacoderma Wieser, 1960 – Nomen nudum Venekey et al. (2014)
P. parabutschlii ( Timm 1961) Theristus (Daptonema) Valid ; spicules>2 a.b.d. Timm (1961) parabutschlii Timm, 1961
P. paranormandica Micoletzky, 1922 Theristus paranormandicus Invalid ; a synonym of Ansari et al. (2013) Micoletzky, 1922; Daptonema normandicum ; Daptonema normandicum spicules only 1.25 a.b.d. (De Man, 1890)
(Continued)
Two opposite and outstretched testes, the anterior one to the left of the intestine, and the posterior one to the right. Spicules paired, slender and arcuate, 79–88 μm long and 4.0–4.4 anal body diameter. Gubernaculum complex, paired large parts slender and proximally pointed, 20–24 μm long; paired small parts stout, distally hook-shaped, pointed in the proximal end, with a ventral apophysis in the middle part, 8–11 μm long. No precloacal supplements ( Figure 2D View Figure 2 ).
Females. Similar to males, but the body slightly larger (1063–1127 μm versus 933– 1023 μm), amphids slightly smaller in both diameter (5.0–5.3 μm versus 6.0–7.3 μm) and corresponding body diameter (23–28% versus 33–46%), buccal cavity broader (10 μm versus 8 μm), head slightly larger (15–18 μm versus 13–16 μm in diameter), and cervical setae sparser ( Figure 2C View Figure 2 ). A single anteriorly outstretched ovary to the left of intestine in three-quarters of specimens, and the rest to the right. Some eggs present in the ovary, the largest ones oblong with narrowly rounded anterior end, up to 97 μm long; the smallest ones roundish, about 4 μm across. Many small circular spermatozoa in uterus, about 2 μm in diameter. Vulva located at posterior third of the body ( Figure 2A View Figure 2 ).
Comparison with related species and genera and overview of Paramonohystera species Paramonohystera was first established by Steiner (1916) as a subgenus of Monhystera Bastian, 1865 being characterized by retractable head, bubble amphids and simple conical buccal cavity, with Monhystera (Paramonohystera) megacephala Steiner, 1916 as the type species. Soon afterward, it was raised to genus level by Filipjev (1918). It is worthy of note that the similar name Paramonhystera as used by Filipjev (1918) and many subsequent authors is an invalid emendation, as clearly stated by Gerlach and Riemann (1973). Two subgenera have previously been proposed for Paramonohystera : Paramonohystera and Leptogastrella ( Wieser 1956; Gerlach and Riemann 1973; Lorenzen 1994). Wieser (1956) defined the two subgenera mainly by the arrangement of cephalic (10 cephalic setae, without additional cephalic setae versus 10–12 cephalic setae, plus additional cephalic setae) and cervical setae (of equal length versus one circle much longer). However, these differences are not distinct and the characters may overlap in a single species (e.g. P. riemanni ; Table 4). On the other hand, all species assigned to the subgenus Leptogastrella have either been synonymized or considered as unreliable reports. Hence, the two subgenera have rarely been employed. The redescription of P. (Leptogastrella) pellucida ( Cobb, 1920) by Wieser (1956) is probably involved in a species complex composed of specimens with about 20 cephalic setae and specimens with a normal circle of 10–12 setae. These specimens might be classified if other typical specimens are available, as stated by Wieser (1956).
Paramonohystera sinica sp. nov. possesses 12 cephalic setae, whereas most known species of Paramonohystera have 10 cephalic setae ( Table 4). As the number of cephalic setae is very stable within species, it seems reasonable to split Paramonohystera and erect a new genus with 12 cephalic setae. However, variability in the number of cephalic setae is usual in Xyalidae , in which Xyala and Cobbia have 10 or 12 cephalic setae, and Theristus and Daptonema have even 10, 12 or 14 cephalic setae. Wieser (1956) also emphasized the variability in the number of the cephalic setae in Paramonohystera . Hence, it is too early to create a new genus before further materials and molecular proofs are available. Within the genus Paramonohystera , only P. buetschlii (Bresslau and Schuurmans Stekhoven in Schuurmans Stekhoven, 1935) , P. concinna Lorenzen, 1977 , P. halerba Fadeeva and Belogurov, 1987 and P. pilosa Boucher, 1971 possess 12 cephalic setae ( Table 4). The new species P. sinica differs from P. buetschlii by body length (933–1023 μm versus 2000–2200 μm) and the ratio of spicule length to a.b.d. (4.0–4.4 versus 2.7). Paramonohystera concinna has segmented cephalic setae which are smooth in P.sinica . Paramonohystera pilosa has much longer spicules (167 µm versus 79–88 µm), broader head (32 µm versus 13– 16 µm) and smaller ratio of spicule length to anal body diameter (2.7 versus 4.0–4.4). In addition P. halerba has two rows of setae on the ventral side of the tail (versus absent in P. sinica ), shorter cervical setae (4–6 μm versus up to 14 μm), longer spicules (105–113 μm versus 79–88 μm) and smaller ratio of spicule length to anal body diameter (2.5 versus 4.0–4.4). All other species of Paramonohystera are easily distinguished from the new species by the number of cephalic setae.
Within the family Xyalidae , Paramonohystera Steiner, 1916 is similar to Daptonema , differentiated by the elongate (>2 a.b.d.) and slender spicules ( Lorenzen 1977; Warwick et al. 1998). Chen and Vincx (2000) recognized nine species of Paramonohystera and provided a key of species including P. breviseta Juario, 1974 , P. longicaudata Timm, 1963 and P. wieseri Ott, 1977 . Among these, P. breviseta has been regarded as a member of the genus Retrotheristus Lorenzen, 1977 . Paramonohystera longicaudata and P. wieseri have relatively short spicules (<1.5 a. b.d.) and probably belong to the genus Daptonema ( Table 3). Chen and Vincx (2000) did not include P. buetschlii (Bresslau and Schuurmans Stekhoven in Schuurmans Stekhoven, 1935) , P. parabutschlii ( Timm, 1961) , P. riemanni ( Platt, 1973) and P. zizichi Pastor de Ward, 1985 etc., without any comments. All of these species have spicules>2 a.b.d., match Paramonohystera well and should be considered as valid species of the genus ( Table 3). Venekey et al. (2014) recognized 18 species of Paramonohystera as valid, and regarded P. micramphis as species inquirenda and P. mystacoderma as nomen nudum. Among the valid species of Venekey et al. (2014), P. longicaudata is probably a member of Daptonema ; P. paranormandica has already been transferred to Daptonema ( Ansari et al. 2013) ; P. pellucida is likely to be a species complex, as mentioned above; P. stricta is probably a member of Promonhystera possessing distinct labial setae; and P. tschilenkoi might be a species inquirenda since the number of cephalic setae that distinguishes Paramonohystera from Retrotheristus was not included in the original description of Platonova (1971), Venekey et al. (2014) also overlooked P. buetschlii , which has been considered as a valid species ( Warwick et al. 1998). Pastor de Ward (1985) previously described a population under the name of Paramonohystera (P.) parabutschlii Timm, 1961 . It is likely a misidentification because the specimens are distinctly longer (2100 μm vs. 918 μm in male) and has a higher number of cephalic setae (12 vs. 10) than the type specimen described by Timm (1961).
Based on the evaluation of 28 nominal species of Paramonohystera , we recognize 14 valid species and provide a tabular key to the genus ( Tables 3, 4). Additionally, we provide an emended generic diagnosis for Paramonohystera : Xyalidae with 6 labial papillae and 10 or 12 cephalic setae usually in six groups, unarmed conical buccal cavity with domed anterior end, circular (mostly) or elliptical amphids, elongate (≥ 2 a.b.d.) spicules, and conico-cylindrical tail with terminal setae. Paramonohystera is most similar to Daptonema and Promonhystera , but differs from Daptonema by the elongate (≥ 2 a.b.d. versus ca.1 a.b.d) and slender spicules and from Promonhystera by the lack of long and distinct labial setae ( Warwick et al. 1998; Coomans and Abebe 2006).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |