
Parmeosaurus scutatus, KEQIN & NORELL, 2000
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2000)249<0001:TCASOL>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC87AC-FFF1-FFD0-99AF-F1BDA053FA91 |
|
treatment provided by |
Felipe |
|
scientific name |
Parmeosaurus scutatus |
| status |
new genus and species |
FAMILY INCERTAE SEDIS Parmeosaurus scutatus , new genus and species
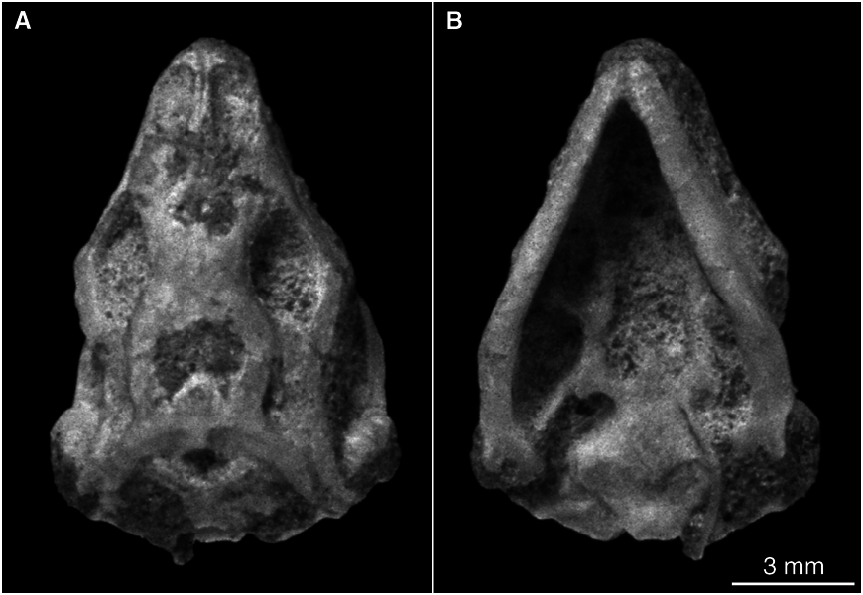
Figures 24 View Fig , 25 View Fig
ETYMOLOGY: parme (Gr.), small light shield; scutatus (L.), armored with shield.
HOLOTYPE: IGM 3 /138 (MAE 9530),
wellpreserved skull with articulated postcranial skeleton.
TYPE LOCALITY AND HORIZON: Ukhaa Tolgod, Mongolian Gobi Desert; Upper Cretaceous Djadokhta Formation (Loope et al., 1998).
KNOWN DISTRIBUTION: Known only from the type locality and horizon.
DIAGNOSIS: Sharing with scincoids the following derived character states: presence of both dorsal and ventral body osteoderms; cephalic osteoderms present; palpebral ossification present; lateral coronoid process of dentary large, extending dorsally onto anterolateral surface of coronoid; retroarticular process inflected medially with small flange on medial margin. Differing from other scincoids in having the following autapomorphies: skull narrow and elongate, with laterally compressed snout; parietal table having welldeveloped lateral flange for dorsal origin of temporal musculature; marginal teeth stout with tricuspid crowns; autotomy fracture present behind caudal ribs.
REFERRED SPECIMENS: IGM 3/139–3/142 (MAE 475/93152, 9627, 9570, 9416), all incomplete skull with mandibles; IGM 3/143 (MAE 9353), fragmentary cranial and postcranial skeleton; IGM 3/144 (MAE 9451), articulated vertebrae with body osteoderms. All the referred specimens are topotypic.
DESCRIPTION
SKULL ROOF: The skull of the holotype specimen is concealed beneath the tail, and is exposed only in ventral view (fig. 24B). However, several other specimens (IGM 3/ 139–3/142) show that the dorsal aspect of the skull is covered with platelike osteoderms. The skull is elongate and the snout is laterally compressed. Anteriorly, the premaxillae are fused and have a slender dorsal spine that deeply intervenes the paired nasals. The nasals have an anterior extension lateral to the premaxillary spine, and posteriorly end with a thin plate that overlaps the frontal.
The paired frontals anterolaterally contact the dorsal process of the maxilla, and this contact separates the prefrontal from the nasal. Posteriorly, the frontals are overlapped by a squared off parietal tab, so that the frontoparietal suture is not transverse (fig. 25A, D). The frontals have parallel lateral borders, showing no interorbital constriction. Along the lateral borders is developed a palpebral series roofing the medial part of the orbit. The frontals have welldeveloped subolfactory flanges, but have no midline contact below the olfactory tract. The descending processes extend anteroventrally, but do not contact the palatines to prevent the prefrontal from entering the orbitonasal fenestra.
The parietal table is penetrated by a small parietal foramen, which is located slightly anterior to the center of the table. At the frontoparietal suture, parietal tabs are present as small triangular structures that are interlocked with the frontals. Laterally, the parietal has a welldeveloped flange, the sloped surface of which indicates a dorsal origin of the temporal musculature. The ventral edge of the flange contacts the prootic bone (best shown on IGM 3/140), but lacks a pointed downgrowth (ventral process of the parietal) that normally contacts the epipterygoid. The supratemporal process of the parietal is elongate, and has an extensive lateral surface for muscle attachment as the extension of the parietal flange. The posterior end of the process reaches the paroccipital process, and apparently has no contact with the quadrate. A small part of the right supratemporal bone is preserved on IGM 3/140.
In lateral view, the maxilla is roughly a triangular bone with the apex of the dorsal process located at the midlevel of the tooth row. The dorsal process contacts the frontal, separating the nasal from the prefrontal. The lateral surface of the maxilla is largely vertical and slightly concave, having six lateral superior alveolar foramina along the ventral border. The posterior maxillary process is dorsally articulated with the welldeveloped lacrimal and jugal, and the latter two elements form the entire ventral border of the orbit. The jugal forms a complete postorbital bar, although its dorsal process is much more slender than the anteroventral process. The element is generally hockeystick shaped, having no trace of a posteroventral process (fig. 25F).
The prefrontal is dorsally covered by the palpebral bones (IGM 3/139). When exposed (IGM 3/140), it shows a welldefined frontal process that extends to the midlevel of the orbit along the lateral border of the frontal. The anteroventral process of the prefrontal forms the entire anterior wall of the orbit and ventrally contacts the palatine. This process laterally contacts the lacrimal and borders the single lacrimal foramen. The process medially enters the orbitonasal fenestra, although the welldeveloped descending process of the frontal forms part of border of the fenestra.
The postfrontal and postorbital are incompletely preserved on IGM 3/140, and are dorsally covered with osteoderms. On IGM 3/ 142, however, the two elements are exposed and seem to be separate as a suture can be identified. The postfrontal is medially forked to clasp the frontoparietal suture, and the postorbital has a long posterior extension forming large part of the supratemporal arch. The supratemporal fenestra is normally developed, although the squamosal is not preserved.
PALATAL ELEMENTS: The vomers are incompletely preserved on several specimens, and are apparently fused with a prominent midline ridge. The posterior vomer extension reaches the level slightly posterior to the midlevel of the tooth row, where the vomers enter the interpterygoid vacuity between the palatines. The palatines are widened and have a strongly concave ventral surface lateral to the vomers. A row of palatal teeth is developed along the medial border of the bone. The posterior border of the palatine is notched for articulation with the pterygoid.
The pterygoid bears two rows of prominent teeth (best shown on IGM 3/140), which are closely packed along the medial border of the palatal process of the bone (fig. 25E). The anterior process of the pterygoid forms large part of the medial border of the interpterygoid vacuity, and apparently has no contact with the vomer. Medially, a welldefined ‘‘mesopterygoid process’’ functions to enhance the articulation with the basipterygoid process of the basisphenoid. The lateral process of the pterygoid is thin but strongly widened, with its posterior border forming a weak crest for the attachment of the pterygomandibularis muscle. Posteriorly, the quadrate process is medially concave, forming a shallow trough.
The ectopterygoid forms part of the posterior border of the suborbital fenestra and has an interlocking articulation with the pterygoid. It has a robust anterolateral extension to form the lateral border of the suborbital fenestra, but whether the extension contacted the palatine cannot be determined given the condition of preservation of the specimens.
BRAINCASE: The braincase floor is elongate, carrying a pit anteriorly and a knob posteriorly, and in the middle of these two structure is a small foramen (best shown on IGM 3/140; see fig. 25E). The basisphenoid and the basioccipital are completely fused, and no suture can be delimited. Anterolaterally, the basipterygoid process is short, and more anteriorly than laterally directed. Between the basipterygoid processes, the rostrum is well ossified and proportionally robust. It extends anteriorly to the same level as the tip of the basipterygoid processes. The braincase floor is posterolaterally expanded, and the sphenooccipital tubercles project ventrally.
Dorsolateral to the floor, a troughlike recessus vena jugularis is deep with the welldeveloped crista prootica forming the lateral wall. Within the recess, the posterior opening of the Vidian canal is located close to the middle of the trough, a short distance from the base of the basipterygoid process. The canal opens at the suture between the fused basisphenoidbasioccipital and the prootic. A much smaller opening (facial foramen) is more posterodorsally located in the trough Laterally at the base of the sphenooccipital tubercle, the occipital recess is small oblique, and narrowly elongate. It does not extend onto the lateral surface of the tubercle. The recess is separated by a small ridge from the more dorsally positioned foramen ovale, which is at about the same level as, but directly posterior to the facial foramen.
The lateral wall of the braincase is observed on the left side on IGM 3/140, which also shows the best view of the braincase floor as described above. As in other lizards the lateral wall of the braincase is mainly formed by the prootic. A welldefined alar process projects anterodorsally contacting the ventral edge of the parietal flange as preserved. Whether the process contacted the epipterygoid cannot be determined, as the latter element is not preserved. Without distortion, the welldeveloped anterior inferior process of the prootic is anteromedially directed lying in a different vertical plane from the alar process. Between the two processes the trigeminal notch is narrow and deep notching to the level of the anterior ampulla
MANDIBLE: The mandible is massive. The posterior border of the dentary has a single surangular notch. Dorsal to the notch, the dentary displays a prominent coronoid process extending onto the anterior surface of the coronoid. Ventral to the surangular is a posteroventral process of the dentary that extends roughly to the same level as the coronoid eminence. The surangular has a strong anterior process fitting into the corresponding notch on the dentary. The anterior surangular foramen opens far posterior from the surangulardentary suture and slightly posterior to the coronoid eminence (IGM 3/139 shows double openings, whereas the holotype and IGM 3/140 show only single openings). The posterior surangular foramen is small and is in the typical position close to the jaw joint The angular is a narrow band, covering the surangularprearticular suture and forming a large part of the ventral border of the postdentary jaw. It has a slender anterior process that intervenes the surangular and posteroventral process of the dentary.
In medial view, the welldeveloped splenial extends anteriorly almost to the symphy sis. The anterior inferior alveolar foramen and the anterior mylohyoid foramen are located posterior to the midlevel of the tooth row, and are located far apart. The posterior extension of the splenial terminates at the level ventral to the dorsal summit of the coronoid. The prearticular and articular are fused. The retroarticular process is broad, slightly inflected medially, and bears a small tubercle (flange) on the medial margin (best shown on IGM 3/139). The mandibular fossa is elongate, opening between the posterior medial process of the coronoid and the craniomandibular joint. The fossa is not expanded or inflated, a condition differing from that seen in extant lacertiforms (Estes et al., 1988).
DENTITION: The marginal teeth are fully pleurodont, having slightly over half of the tooth height attached to the lateral parapet of the tooth row. The premaxillae have as many as nine teeth, and these are unicuspid and are about equal in size. The maxillary tooth row contains 17–18 teeth, and the dentary contains roughly 22 teeth. The first four or five teeth on both maxilla and dentary are relatively slender and unicuspid, those on the middle part of the tooth row have stout crowns and are bicuspid with a small anterior accessory cuspule. The most posterior teeth are shortcrowned, very stout, and weakly tricuspid.
POSTCRANIAL SKELETON: The postcranial skeleton is best preserved on the holotype. The actual number of its presacral vertebrae cannot be determined, but it appears to have had 26 or more (in keeping with the elongation of the body). The vertebrae are slightly elongate, with procoelous centra and low neural spines. The centra are procoelous. Zygosphenes and zygantra are absent, and therefore, there are no accessory articulations between vertebrae. The caudals are more strongly elongate than the presacrals and have a longitudinal furrow on the ventral surface of the centrum. At least the anterior caudals have a single pair of caudal ribs fused to the centrum. The autotomy fracture is developed right behind the caudal ribs—a condition known for some iguanians and gekkotans (Estes et al., 1988).
The pectoral and pelvic girdles on the holotype cannot be exposed without damaging the covering sheet of osteoderms, but some long bones can be observed in ventral view. The left forelimb has the radius exposed as a slender element. The right hindlimb is well preserved, having the femur, tibia, and a metatarsal preserved in articulation. The hindlimb is quite robust, and the tibia is slightly shorter than the femur (fig. 24B).
OSTEODERMS: One of the notable features of this lizard is the presence of osteoderms covering both dorsal and ventral aspects of the skull and the body. The osteoderms on the skull roof are thin and platelike with essentially no imbrication. Those covering the ventral aspect of the skull are significantly smaller than those on the skull roof, and are more or less rhomboid and imbricated.
The dorsal body osteoderms are rectangular. Having a welldeveloped gilding surface anteriorly, these are imbricated one another anteroposteriorly but are sutured laterally. The ventral osteoderms are also rectangular, but are even smaller than the dorsal ones. Each individual osteoderm is about half the size of those covering the dorsal side of the body; therefore, there are more ventral rows of osteoderms than dorsal ones. All the osteoderms have smooth surfaces and are not keeled.
COMPARISON AND DISCUSSION
The referral of Parmeosaurus scutatus to the Scincomorpha is supported by two character states: nasalprefrontal contact is lost, owing to the presence of a frontalmaxillary contact; and the lateral process of the coronoid is overlapped anteriorly by the dorsal process of the dentary, so that the lateral exposure of the coronoid process is limited to a narrow wedge between the dentary and surangular (see Estes et al., 1988 for evaluation of the characters). Presence of both dorsal and ventral body osteoderms merits further investigation: it occurs in Scincidae and Cordylidae , as well as the Paramacellodidae (Evans and Chure, 1998) . The rectangular osteoderms in this Cretaceous lizard are similar to cordylids and paramacellodids, but differ from the cycloidtype in most skinks. This character was not investigated in either Estes et al. (1988) or Presch (1988) and polarity is yet to be established.
Within the Scincomorpha, the new species cannot be reliably classified in a particular subgroup because of conflicting evidence. It shares with the Scincoidea ( Scincidae + Cordylidae ) several derived character states. These include: cephalic osteoderms present; lateral coronoid process of dentary is large, extending dorsally onto anterolateral surface of coronoid; retroarticular process is inflected medially; medial margin of the retroarticular process has a tubercle or a small flange; retroarticular process is broadened posteriorly (see Estes et al., 1988 for character evaluation). In contrast, at least two character states indicate a possible lacertiform relationship of Parmeosaurus scutatus : presence of a pterygoid lappet of the quadrate (best shown on IGM 3/139), and a facial region that is elongate with the snout laterally compressed (see Estes et al., 1988). Another character state, presence of palpebral ossifications, has been interpreted as a separate synapomorphy for the Scincoidea, Lacertidae , and the Anguimorpha (see Estes et al., 1988), but it also occurs in the fossil group Paramacellodidae , which is probably the sister taxon of the Scincoidea (Evans and Chure, 1998). Therefore, this character state should be interpreted as a synapomorphy for a more inclusive group. Alternatively, if the Paramacellodidae represent the basal clade of the Scincomorpha (a hypothesis that needs to be tested), it would be a synapomorphy of Autarchoglossa with secondary loss within the Scincomorpha.
The available evidence seems to support referral of the new species to the Scincoidea, but conflicting evidence, such as having a pterygoid lappet on the quadrate and facial elongation, significantly weakens this referral. Pending wholesale revision of related taxa and the complete description and preparation of additional material, we tentatively refer Parmeosaurus scutatus to the Scincoidea.
Hymenosaurus clarki , new genus and species Figure 26 View Fig
ETYMOLOGY: hymen (Gr., god of marriage) + sauros (Gr., lizard), a present for Jim Clark’s wedding; the species epithet is to rec ognize Clark’s profound contribution in collecting vertebrate fossils, including many lizard specimens, from the Gobi Desert.
HOLOTYPE: IGM 3 /53 (MAE 154/9340), incomplete skull with mandibles.
TYPE LOCALITY AND HORIZON: Ukhaa Tolgod, Nemegt Basin, Mongolian Gobi Desert Upper Cretaceous Djadokhta Formation (Loope et al., 1998).
DIAGNOSIS: Sharing with scincoids character states such as: retroarticular process inflected medially, broadened posteriorly; sharing with scincids presence of jugalsquamosal contact.
Differing from other scincomorphs in having the following derived character states presence of moderate retraction of narial openings; frontals proportionally wide; presence of prefrontalpostfrontal contact; parietal foramen greatly reduced as minute opening and located close to posterior border of parietal table; occipital condyle formed exclusively by basioccipital; posterior notch of dentary articulates with surangular only, without involvement of angular; retroarticular process posteriorly notched; sphenooccipital tubercle greatly reduced, and shifted anteriorly close to midlevel of braincase floor.
DESCRIPTION
The entire skull and the mandibles are generally well preserved without distortion However, the dorsal surface of the skull roof was exposed in the field and the elements such as the nasals and the frontals are eroded and incomplete.
SKULL ROOF: The premaxillae are clearly paired, with extremely slender and elongate dorsal spines. The nasals are incompletely preserved on both sides. Although the midline suture is not shown as preserved, there is no indication of fusion of the two bones.
The frontals are clearly separated by a midline suture and are proportionally wide with parallel lateral borders. Although lacking a slender anterolateral process, the frontal contacts the dorsal process of the maxilla, so that the nasals are separated from the prefrontals. The frontals have parallel lateral borders, lacking welldefined interorbital constriction. The lateral border is also ex cluded from the medial rim of the orbit, because of the abutting contact of the prefrontal with the postfrontal. Ventrally, the subolfactory flange is fairly well developed, but is relatively shallow and has no contact along the midline.
The parietal table is roughly rectangular in shape. It lacks lateral flanges, indicating a ventral origin of the temporal muscles; however, the posterior border of the table has a welldeveloped flange for attachment of the axial muscles. The parietal foramen is greatly reduced to a minute opening lying close to the posterior border of the parietal table (fig. 26A). Like in many extant skinks, the supratemporal process of the parietal distinctly bends so the basal part of the process extends laterally, then turns more posteriorly. Distally, this process contacts the paroccipital process and is separated from the quadrate by the supratemporal and the squamosal.
The prefrontals have a slender and elongate frontal process extending posteriorly to contact the anterior process of the postfrontal. This contact excludes the frontals from the medial rim of the orbit. The postfrontal and postorbital are clearly separate elements. The postfrontal is proportionally larger than the postorbital, and is medially forked to clasp the frontoparietal suture. It has an elongate anterior process contacting the prefrontal, and the two bones together form the medial rim of the orbit and exclude the frontal from entering the orbit.
The squamosal is preserved on both sides. The element lacks a dorsal process, but the anterior process is strongly elongated to have an abutting contact with the jugal. The supratemporal is clearly identifiable on the right side of the specimen. It medially attaches to the lateral surface of the paroccipital process and the supratemporal process of the parietal, and laterally contacts the squamosal and the cephalic head of the quadrate. The quadrate is strongly widened as commonly seen in extant skinks and lacks a pterygoid lappet.
The maxilla is completely preserved on both sides. The lateral surface of the bone is smooth without osteodermal ornamentation. Above the ventral border is a shallow groove, in which opens a horizontal row of lateral superior alveolar foramina. Anteriorly, the premaxillary process is well developed in keeping with the narial retraction. It has a sutural articulation with the premaxilla, leaving no aperture between the two elements. The anteromedial process is short, showing no tendency to develop a midline contact with its opposite element. In keeping with the moderate narial retraction, the dorsal process of the maxilla rises above the middle level of the tooth row. The process is roughly triangular, and bends medially to form a facial ‘‘table’’ together with the nasal and prefrontal. The posterior process forms the lateral part of the ventral border of the orbit, as the jugal contributes to the medial part of the border.
The lacrimal is apparently absent, as it cannot be identified on either side. The jugal is generally boomerangshaped. At the posteroventral corner it has a blunt, but prominent, process directed ventrally. The posterodorsal process is equilateral along its entire length without the distal narrowing seen generally in other lizards. It has a blunt dorsal tip which contacts both the postorbital and the squamosal. The anteroventral process is attached to the medial side of the posterior process of the maxilla, so that the jugal cannot be seen in lateral view.
PALATAL ELEMENTS: The palatal elements are mostly well preserved. The vomers are short and seem to be paired as preserved. The palatal surface of the element is toothless. The palatine is short and wide, but it lacks the characteristic scrolling seen in skinks and has no midline contact with the opposite side of the element. The palatine has a posterior process, which contacts the pterygoid and forms most of the medial bor der of the suborbital fenestra. No palatal teeth are developed on the ventral surface of the element.
The pterygoid is a typical Y shaped bone, with short anterior processes and a longer posterior process. The anteromedial process is slender, but small denticles are identifiable on the ventral surface. The anterolateral process forms the entire posterior rim of the suborbital fenestra, and contacts the ectopterygoid to form a very weak ventral process (pterygoid process). The ectopterygoid is small, and its short anterior process forms the posterior half of the lateral border of the suborbital fenestra. Thus, the maxilla contributes to the anterior half of the border of the suborbital fenestra.
BRAINCASE: The braincase floor is well preserved, and is slightly convex ventrally The basioccipital and the basisphenoid are fused, as no suture can be delimited between the two elements. Like in many extant scincoids, the floor is narrow at the base of the basipterygoid processes, strongly widened at the level of the sphenooccipital tubercles and narrow posteriorly at the base of the occipital condyle (fig. 26B). The basipterygoid process is short and laterally directed. The cultriform process has a short but wellossified base. The sphenooccipital tubercle is greatly reduced as an extremely small knob and is shifted anteriorly close to the midlevel of the braincase floor. The occipital recess is a fissurelike structure and is strongly oblique anteroventralposterodorsally. The occipital condyle is small, and is formed entirely by the basioccipital, with essentially no contribution from the exoccipitals. The articulation of the exoccipital with the basioccipital is marked by a ridge, indicating possible fusion of the two bones.
The recessus vena jugularis is greatly reduced as an extremely shallow groove. The crista prootica is a low crest, in keeping with the reduction of the recessus vena jugularis The posterior opening of the Vidian canal is very small and is located at the base of the basipterygoid process. Posteriorly within the recessus, the facial foramen opens on the medial wall of the groove and just anterior to the knoblike sphenooccipital tubercle. The foramen ovale is slightly elongate and is located right above the oblique occipital recess
In dorsal view, the entire braincase is shift ed posteriorly and well exposed posterior to the parietal margin (perhaps a burrowing adaptation, see Rieppel, 1981). In occipital view, the paroccipital process is short and is proportionally robust. It is horizontal in pos terior view, but is anterolaterally directed in dorsal view, differing from the normal condition in which the process is posterolaterally oriented. This feature of the paroccipital process, seen in many extant skinks, is correlat ed with the posterior shifting of the braincase as a burrowing adaptation.
MANDIBLE: The mandible is extremely slender. In lateral view, the dentary is penetrated by five extremely small mental foramina. The posterior border of the dentary is slightly notched for the blunt anterior process of the surangular. The surangular is the only element that fits in the dentary notch, and the anterior part is separated from the angular bone by a slender posterior ventral process of the dentary. This condition is similar to that seen in typical burrowing skinks (e.g., AMNH 2245: Scincus officinalis ; AMNH 48509: Acontias gigracilicauda ), but differs from other scincomorphs in which normally both the surangular and angular fit in the dentary notch.
In medial view, the subdental shelf is slen der but clearly defined, unlike the sloped toothbearing border in advanced anguimorphs ( Anguidae and Platynota). The Meckelian canal is narrow and largely covered by the splenial. The splenial is narrowly elongate, with its anterior extension terminating close to the mandibular symphysis and its posterior extension terminating below the posteroventral process of the coronoid bone. The anterior inferior alveolar foramen and the anterior mylohyoid foramen cannot be properly identified as a matter of preservation.
The retroarticular process is medially inflected and posteriorly widened. The most distinctive feature of the lower jaw is the posterior notch of the retroarticular process. This notch may be functionally related to attachment of the mandibular depressor muscles, but its actual role is unclear. To our knowledge, no other lizard known has developed this type of notch.
DENTITION: Marginal teeth are preserved on both sides of the upper and lower jaws, but neither side of the jaws shows a complete tooth row. The teeth are slenderly cylindrical, and closely spaced from one another along the tooth row. Tooth implantation is pleurodont, as about half of the tooth height atta ches to the relatively high lateral parapet of the tooth row. The crowns are poorly preserved, the cusp pattern cannot be clearly viewed on the specimen.
COMPARISON AND DISCUSSION
The new taxon is possibly referable to the Scincoidea on the basis of the feature retroarticular process inflected medially and broadened posteriorly. Within the Scincoidea, it shares with scincids the presence of a jugalsquamosal contact and expansion of the postfrontal, but lacks any tendency toward developing a secondary palate (see Estes et al., 1988 for character evaluation).
A notable feature of the skull is that the prefrontal contacts the postfrontal above the orbit and along the lateral border of the frontal. Such a contact occurs homoplastically in several groups of lizards, including chamaeleonids, some skinks, and some anguimorphs (Estes et al., 1988). Whether such a condition seen in the new taxon is homologous to the similar condition in those skinks cannot be determined, because of the uncertain relationships of the new taxon within the Scincomorpha.
Generally, the characters discussed above indicate a possible scincoid relationship of this new lizard; however, lack of other character support (see Estes et al., 1988) prevents us from placing it in any of the two extant familial groups of the Scincoidea ( Scincidae and Cordylidae ). Also, further work needs to be done to resolve its relationships with other scincomorphs such as Parmeosaurus , Slavoia , Globaura , and Eoxanta .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.