
Lazulina Willmott, Nakahara & Espeland, 2023
|
publication ID |
https://doi.org/ 10.1111/syen.12590 |
|
publication LSID |
lsid:zoobank.org:pub:1679054D-6E3B-4B80-B8D6-0ED6628ADE81 |
|
DOI |
https://doi.org/10.5281/zenodo.7909471 |
|
persistent identifier |
https://treatment.plazi.org/id/03EC879F-FF9C-FFC9-A8B0-AC5FFDF91201 |
|
treatment provided by |
Julia |
|
scientific name |
Lazulina Willmott, Nakahara & Espeland |
| status |
gen. nov. |
Lazulina Willmott, Nakahara & Espeland , genus novum.
Type species — Euptychia hewitsonii Butler, 1867 , by present designation.
Zoobank registration: https://zoobank.org/Nomenclatural Acts/2F9B7D53-51B3-4A62-B5F1-AE2312CD4163
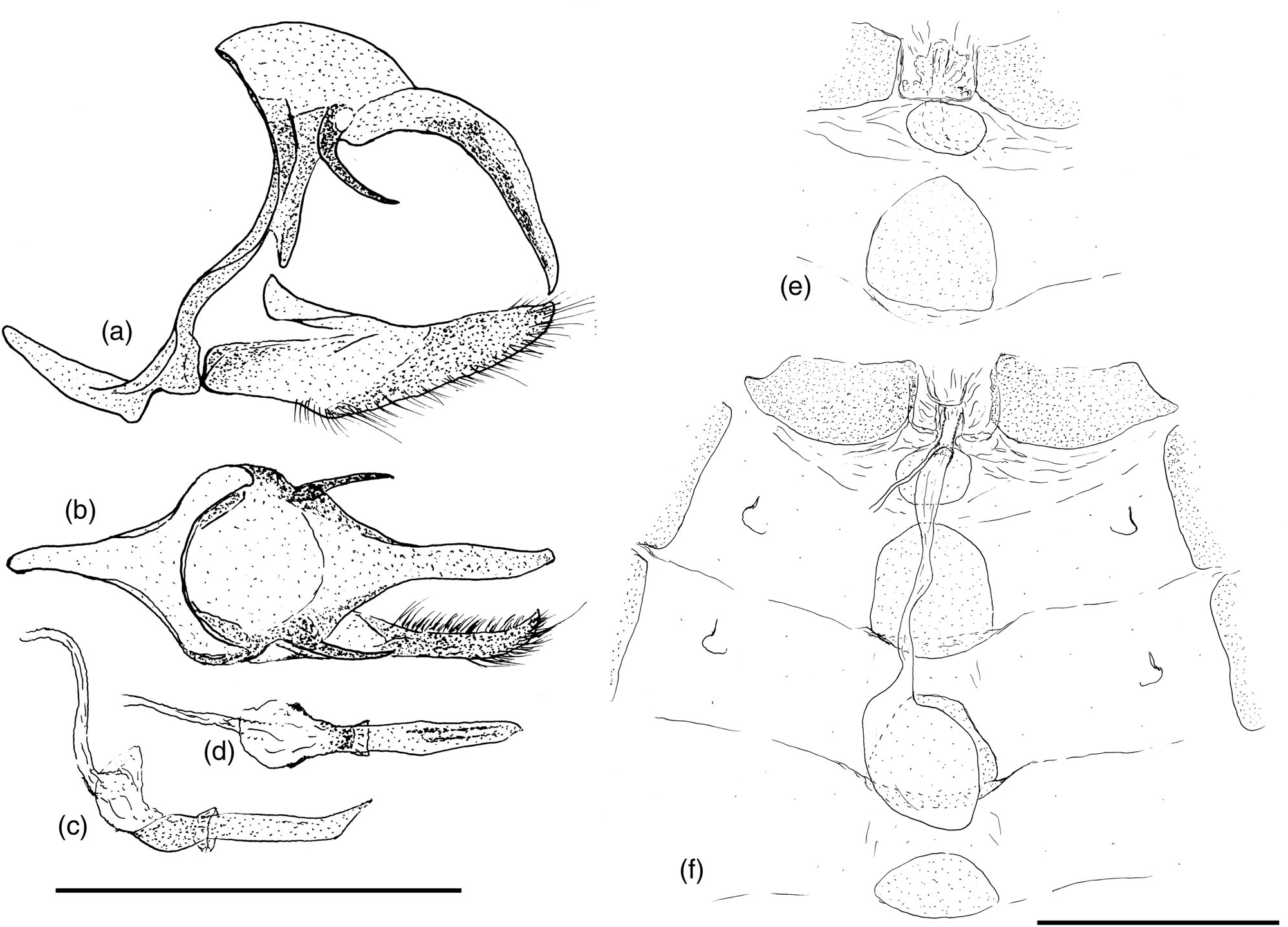
Systematic placement and diagnosis. Lazulina gen.n. is wellsupported as monophyletic (SH-aLRT = 100, UFB = 100), but only moderately supported as sister to Cristalinaia (SH-aLRT = 89.2, UFB = 90), with the clade containing these two genera placed without strong support as sister to the Hermeuptychia clade. The new genus can be distinguished from all other Euptychiina by a rather large number of distinctive possible synapomorphies in the male, including: (a) a dark band of androconial scales just posterior of the cubital vein on the DFW extends from 2A-Cu 2 into Cu 1 -M 3, with associated modified scales ( Figure 11a – c View FIGURE 11 ; see Description below for details). (b) an elongate cluster of long, hair-like scales (‘hair-pencil’) is located near the HW base in cell 3A-2A, the distal tip of which covers an elongate, oval patch of modified scales surrounding vein 2A ( Figure 11a,c View FIGURE 11 ). A more elongate patch of somewhat similar androconial scales is present in HW cell 2A-Cu 2 in Atlanteuptychia ( Freitas et al., 2013) . (c) the basal half of HW vein 2A is bowed outwards, towards the adjacent ‘hair-pencil’ ( Figure 11 View FIGURE 11 ). (d) Fourth and fifth sternites of the male abdomen are reduced to a small, sclerotized band at the anterior edge of each segment, with the remainder of each segment forming a flexible pocket that contains dense black elongate rectangular androconial scales, which clearly stand out beyond the ventral abdominal edge in lateral view ( Figure 11g View FIGURE 11 ). Similar scales are present dorsally on the terminal two abdominal segments of Atlanteuptychia ( Freitas et al., 2013) . A further possibly distinctive character is the reduction of the middle of FW vein M 2 -M 3, and numerous other characters distinguish the genus from its possible sister genus, the highly autapomorphic Cristalinaia (see Freitas, Mota, Zacca, & Barbosa, 2019). Overall, the iridescent blue dorsal wing colour of Lazulina gen.n. species readily distinguishes them from most euptychiines, except for females in comparison with several species of Chloreuptychia and Pseudeuptychia . Lazulina gen.n. can be distinguished by the presence of yellowish or reddish brown scaling within the dark band posterior of the VFW apical ocellus, and by the VHW ocellus in M 3 -M 2 being basally displaced in comparison with that in M 2 -M 1.
Etymology. The generic name is a feminine noun in the nominative singular, derived from the Latin adjective ‘lazulinus’, meaning something that is blue, in reference to the distinctive blue dorsal wing markings of this genus.
Description ( Figures 11 View FIGURE 11 and 12 View FIGURE 12 ). Some notable characters include: eyes naked; pterothoracic legs dorsally darker, tibia with two principal longitudinal rows of spines ventrally, pair of spurs of similar length at distal end of tibia, first tarsomere with three principal longitudinal rows of spines ventrally, remaining tarsomeres with four principal longitudinal rows of spines ventrally. Small Euptychiina (FW length typically 17 – 21 mm), FW triangular and somewhat elongate, HW margin not scalloped, slightly angled at end of vein Cu 1 and slightly elongated towards tornus. Sexes dimorphic, dorsal wings dark brown: Male: HW with iridescent blue to purple scaling at least in posterior half, to covering most of wing, L. hewitsonii comb.n. partially translucent pale greyish in HW discal cell and with iridescent blue scaling along anal margin and distal margin of FW; middle of FW with complex configuration of androconial scales, comprising long hair-like scales in posterior third of discal cell, which appear to be ‘swept-back’ towards cubital vein, cubital vein bordered posteriorly by a band of broad, short, rectangular pale brown scales sparsely covered with long hair-like scales, then a band of dense, dark androconial scales that are attenuated towards tip then terminate in a curved, brush-like structure; 8th abdominal tergite reduced dorsally, leaving a sclerotized strip along anterior edge; genitalia with downwardly curving uncus longer than tegumen, brachia short and pointing ventrally of uncus, valva ventral edge distinctly angled in middle, terminating in blunt tip; saccus short (similar in length to tegumen); aedeagus short, anterior portion opening dorso-anteriorly (rather than dorsally) and slightly flared in dorsal view, with two narrow bands of cornuti. Female: lacking androconial scales, with faint purplish blue iridescence over both wings or HW only, not extending to distal margins. Ventral wings with ground colour ranging from brown to silvery grey; dark reddish brown discal and postdiscal lines traversing both wings, discal line angled inwards at HW anal margin, postdiscal line displaced towards tornus in cell 2A-Cu 2; VFW with a single subapical ocellus in cell M 2 -M 1, black with a yellow ring and 1 – 2 silvery elongate pupils (when 2 pupils are present they unite to form a ‘C’-shape), placed within a broad, dark brown postdiscal band (umbra); posterior of ocellus, VFW umbra has dark orange-brown scaling or yellowish brown scaling, in some species bordered basally by a pale purplish band; two dark marginal lines, straight and parallel to wing margin; VHW has five postdiscal ocelli as follows: In Cu 2 -Cu 1, largest ocellus, black with yellow border that extends into adjacent cells, with two elongate silver dashes as pupils; in Cu 1 -M 3, elongate brown ocellus with yellow border that may be absent distally, with large central elongate silver pupil; in M 3 -M 2, similar to preceding but smaller and displaced basally; in M 2 -M 1, black ocellus with yellow ring filling cell, two silver pupils united to form distally pointing ‘C’-shape; in M 1 -Rs, similar to preceding but smaller; ocelli bordered distally by dark brown, especially distal of smaller ocelli, then pale purplish surrounding two dark marginal lines which become gradually thicker and more reddish towards tornus; genitalia with each sternite reduced posteriorly; eighth tergite reduced to small sclerotized posterior patch, intersegmental membrane between seventh and eighth abdominal segments somewhat pleated and expandable with an oval sclerotized plate ventrally, eighth segment with large irregular lateral sclerotized plate, lamella antevaginalis and antrum unsclerotized, ductus bursae narrow and unsclerotized, corpus bursae small, circular and with two narrow signa.
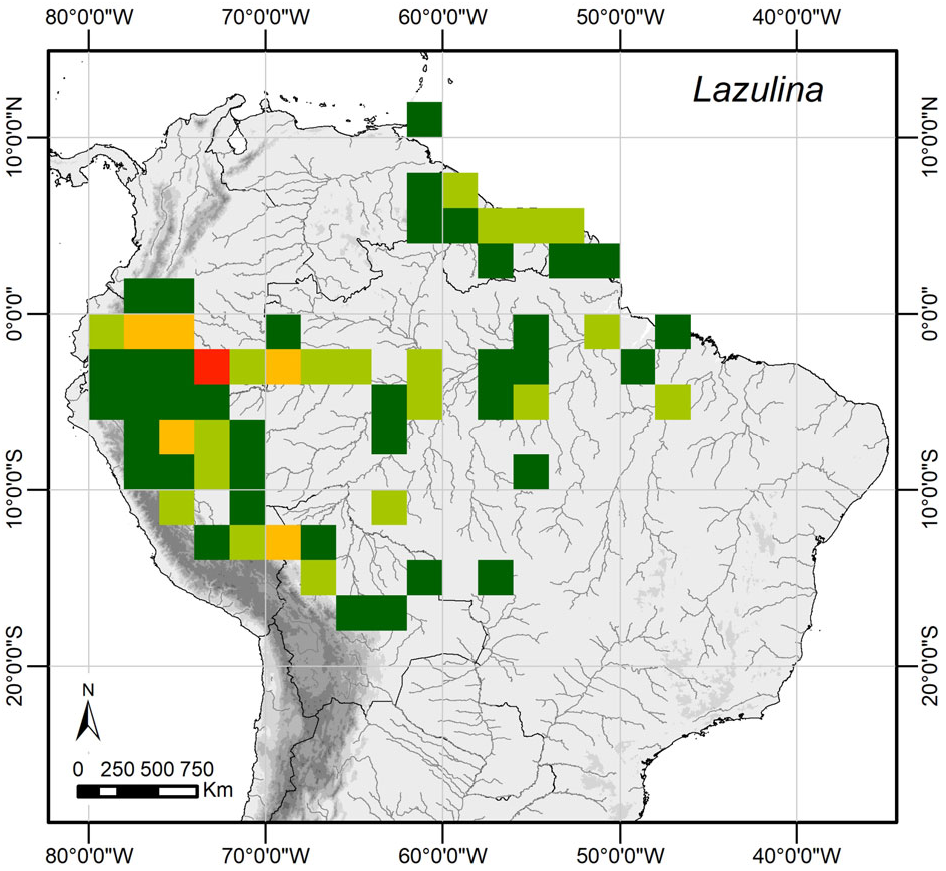
Distribution and natural history ( Figure 13 View FIGURE 13 ). Lazulina gen.n. species occur from sea level to 1200 m in the Amazon, Guianas and Trinidad, with the highest local diversity occurring in the upper Amazon. All species are found in the understory of disturbed to primary rainforest, often near forest edges at rivers, streams or palm swamps, where they typically occur as solitary individuals. There is no published information on the hostplants or immature stages, although some data on the immature stages of L. tolumnia comb.n. are provided in Murray ’ s (2001b) unpublished dissertation.
Discussion. Butler (1867) described the type species for this genus, Euptychia hewitsonii , based on both male and female specimens, from Pará (Brazil), then in the collections of Bates, Salvin, Hewitson and the ‘B[ritish].M[useum].’ (now NHMUK), along with a male or males from Ega (now Tefé, Brazil), then in the last of these collections. The description is detailed, notably mentioning the somewhat translucent basal area of the wings that are distinctive to this species, and the ventral surface of a male was illustrated on pl. 40, Figure 4. A View FIGURE 4 syntype in the NHMUK was examined and matches the original illustration.
The three described species placed here in Lazulina gen.n were placed until now in Chloreuptychia ( Forster, 1964; Lamas, 2004), a genus now split into four genera in four clades (e.g. Nakahara, Lamas, et al., 2019; this publication). However, Weymer ( Weymer, 1911, p. 220) placed the three described Lazulina species in a distinct ‘Tolumnia Group’, correctly noting that males of these species shared a band of androconial scales just posterior of the discal cell on the DFW. Forster (1964, p. 121) used this character in his key to separate out these three species, but nevertheless grouped them with other unrelated ‘blue’ species in his new genus Chloreuptychia . The molecular analysis here shows that Lazulina gen.n. are not closely related to any species formerly included in Chloreuptychia , and indeed the relationships of this highly distinctive genus are still not confidently known. There are a number of remarkably similar structures in Lazulina gen.n. and Atlanteuptychia , including androconial scales in the middle of the DFW, a DHW androconial ‘hair-pencil’, and dense black androconial scales on the male abdomen. Nevertheless, differences in the morphology and position of the androconial structures support independent derivation, the female genitalia and wing patterns show no particular similarity, and molecular data, based on multiple individuals of each genus, indicate that these are not closely related genera, placing them in distinct clades.
Lazulina Willmott, Nakahara & Espeland , gen.n.
catharina (Staudinger, [1886]) , comb.n., was Chloreuptychia hewitsonii ( Butler, 1867) , comb.n., was Chloreuptychia = polla (MÖschler, 1883), comb.n., was Chloreuptychia tolumnia (Cramer, 1777) , comb.n., was Chloreuptychia

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |