
Otiorhynchus ( Arammichnus ) indefinitus Reitter, 1912
|
publication ID |
https://doi.org/10.11646/zootaxa.4108.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B802F2B1-944E-4B84-A856-8091E60D88FC |
|
DOI |
https://doi.org/10.5281/zenodo.6062724 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB857D-FFFF-AD03-0592-FD7342DB1184 |
|
treatment provided by |
Plazi |
|
scientific name |
Otiorhynchus ( Arammichnus ) indefinitus Reitter, 1912 |
| status |
|
Otiorhynchus ( Arammichnus) indefinitus Reitter, 1912 View in CoL
We took over this name from Löbl & Smetana (2012) despite the absence of a formal taxonomical act of synonymization, as we believe that the authors are right.
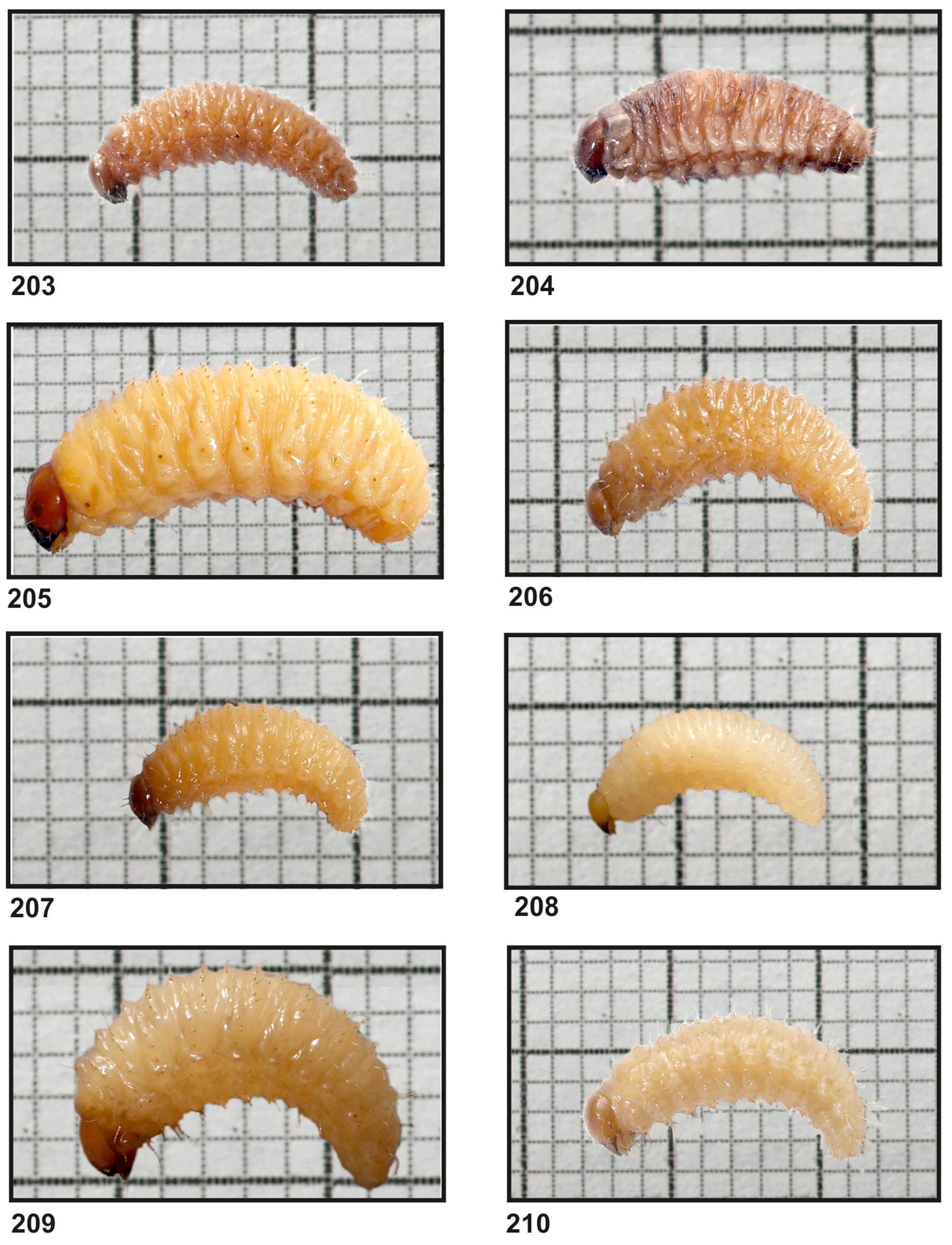
Material examined: 11 larvae ( Fig. 203 View FIGURES 203 – 210 ). JKI, Braunschweig, climate chamber, 21.09.2011, 1 ex.; dto., 28.10.2011, 5 ex.; dto., 0 1.11.2012, 4 ex.; and 1 ex. on 23.04.2013 from two flowerpots that were taken to the Curculio Institute in Hannover in winter 2012 / 2013 for regular pupal search.
Remarks about breeding and development. All larvae were received from breeding efforts in the climate chamber of the JKI in Braunschweig from flowerpots with Ligustrum vulgare L. and Syringa vulgaris L. Breeding attempts were started on 24.06.2011 and on 24.05.2012. In 2011 16 adults and in 2012 8 adults and 70 eggs from Hannover-Herrenhausen, Berggarten, and Hannover-Linden, park area, were placed in 2 small flowerpots with Ligustrum vulgare and 3 flowerpots with Syringa vulgaris in the JKI. In 2012 5 flowerpots with only Syringa vulgaris were used. In the Berggarten, an old botanical garden, the weevils were mainly collected from Dendranthema x grandiflorum (Ramat.) Kitam. cultivars.
The first mature larvae were obtained in 2011 at the end of September and in 2012 at 1st November. Then development was delayed in many specimens. Mature larvae were also present in November, December, January, February, but on 0 1.11.2012 already one recently emerged adult was obtained. The only pupa was detected in June, on 16.06.2012, and described for the first time by Gosik & Sprick (2013). The reasons for this developmental delay are unknown, as other originally Mediterranean species, like Otiorhynchus armadillo , O. aurifer or O. meridionalis , did not show such a delay which is regarded to be an adaptation to lower central European winter temperatures.
The life-cycle of Otiorhynchus indefinitus is not well known, but apparently rather similar to O. raucus (see next species). Teneral adults, usually not sclerotized specimens, appear early in the season on the soil surface. Activity decreases gradually during summer and ends in September. Up to now neither larvae nor pupae were found in the field, but 2 recently emerged adults were recorded in September, indicating that pupation took place in the same year as egg deposition. But it is unknown if larvae also overwinter (originated from eggs deposited later in the season) as in most other Otiorhynchus species with well known life-cycle. In Wiesbaden-Schierstein (South Hesse, Germany), G. Kubach found three not sclerotized, overwintering teneral adults in soil, but less deep than O. raucus ( Sprick & Stüben 2012) . Very likely, these specimens stayed in their pupal chambers. The low age of these specimens can also be detected by the presence of mandible appendages that will be lost just before the first food uptake. The appearance of single specimens at the soil surface in September might be due to a disturbance of the overwintering site, e.g. by agricultural or horticultural activities. Breeding in flowerpots proved to be practicable in this species, and its susceptibility to entomopathogenic nematodes was tested in a project about soil-dwelling weevils (Ufer et al. 2011; cited in Sprick & Stüben 2012).
HW HH BL BH
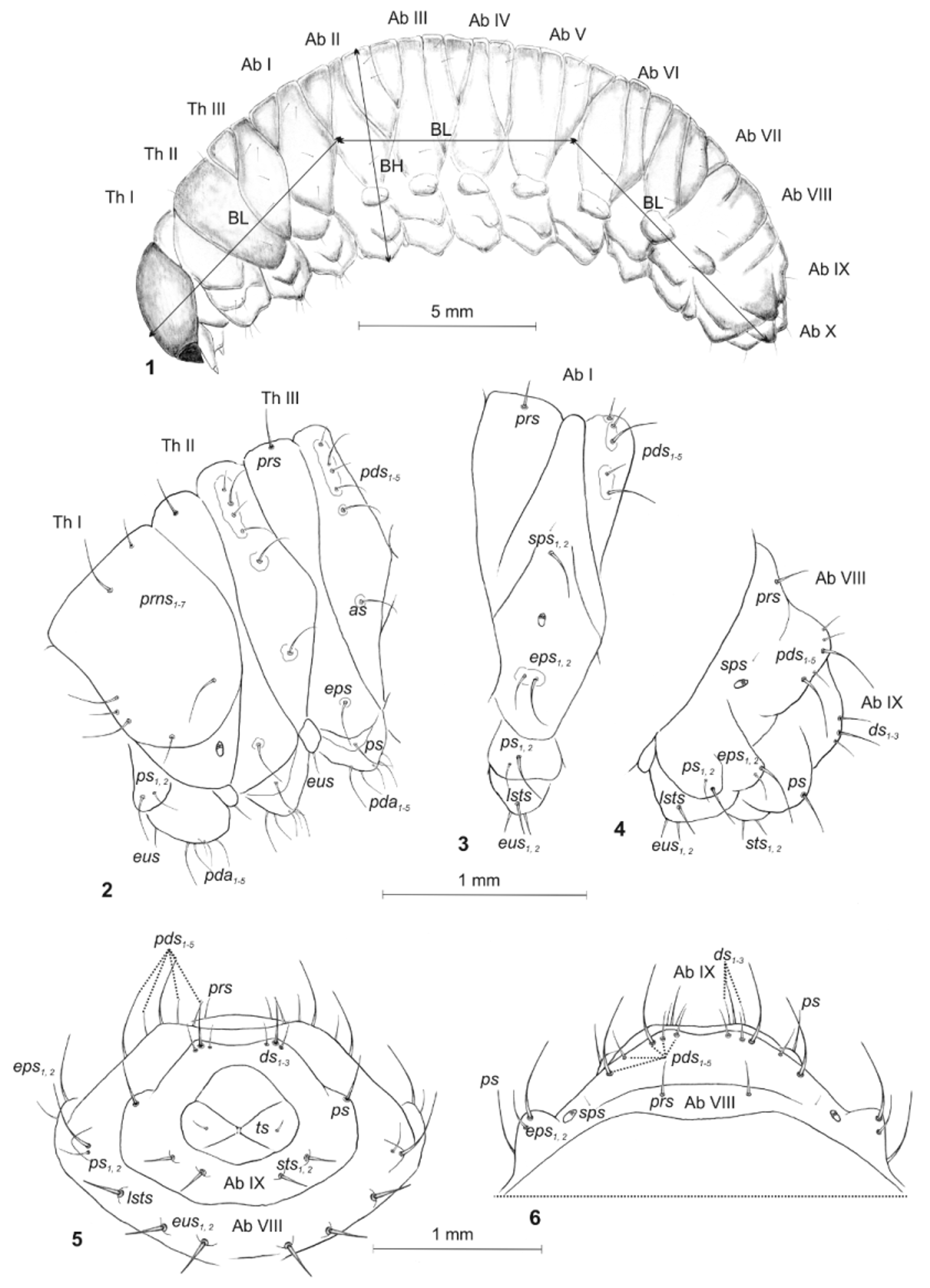
Species min. median. max. min. median. max. min. median. max. min. median. max. O. indefinitus 1.20 1.20 1.40 1.10 1.20 1.30 6.80 7.60 9.00 2.30 3.00 2.40 O . raucus 0.50 0.70 0.70 0.50 0.50 0.80 2.50 2.70 3.10 0.90 1.00 1.40 O . ligustici* 2.40 2.50 3.00 2.40 2.50 2.60 12.00 12.70 13.00 4.00 4.80 5.60 O . porcatus* 0.75 1.15 1.45 0.65 1.00 1.50 4.50 6.00 7.30 1.90 2.20 2.90 O . sulcatus 1.60 1.70 1.90 1.50 1.60 1.80 7.40 9.60 11.70 2.70 3.30 3.80 O . singularis 1.60 1.60 1.70 1.50 1.60 1.80 6.50 7.70 10.10 2.50 3.00 3.50 O . armatus 1.70 1.70 2.00 1.50 1.70 1.80 6.90 8.40 10.40 2.20 3.10 3.50 O . armadillo 1.70 1.70 2.00 1.70 1.80 2.00 8.20 9.10 9.80 3.00 3.20 3.50 O . aurifer 1.70 1.70 1.90 1.70 1.70 1.80 8.20 8.50 10.50 2.50 3.30 3.50 O . meridionalis 1.80 1.90 1.70 1.80 8.00 10.50 3.10 3.10 O . clavipes 2.00 2.10 2.30 1.80 2.00 2.10 7.00 9.20 11.20 3.70 4.0 5.20 O . fuscipes 2.00 2.10 2.40 1.80 2.00 2.10 7.20 9.60 11.60 3.10 4.0 4.90 O . lugdunensis 1.70 1.80 2.20 1.80 1.80 2.20 7.80 9.80 11.20 2.60 3.20 3.80 O . salicicola* 1.55 1.75 1.80 1.20 1.30 1.50 7.50 9.50 10.00 3.00 3.20 3.40 O . ovatus 1.00 1.10 1.20 1.00 1.10 1.20 4.30 6.20 6.70 1.10 2.20 2.30 O . crataegi 1.00 1.10 1.10 1.00 1.00 1.10 6.00 6.30 7.00 1.90 2.00 3.40 O . rugosostriatus 1.50 1.60 1.70 1.40 1.50 1.60 7.70 8.70 9.00 2.30 2.80 3.00 Description ( Figs. 1–11 View FIGURES 1 – 6 View FIGURES 7 – 11 ).
Coloration. Head dark brown; all thoracic and abdominal segments dark yellow brown; cuticle almost smooth. Body rather compact ( Fig. 1 View FIGURES 1 – 6 ). Chaetotaxy. Setae different in length, filiform, greyish. Thorax. Prothorax with 4 long and 3 medium long to short prns; and 2 ps, different in length. Mesothorax with 1 long prs; 5 pds (ordered: 1 short, 1 long, 1 short and 2 long); 1 long as; 1 long eps; and 1 long ps. Chaetotaxy of meso- and metathorax similar. Each pedal area of thoracic segments well isolated, with 5 pda, different in length. Each thoracic segment with 1 short eus ( Fig. 2 View FIGURES 1 – 6 ). Abdomen. Abdominal segments I–VIII with 1 short prs; 5 pds (ordered: 2 short, 1 long, 1 short and 1 long); 1 long and 1 minute sps [abd. seg. VIII only with 1 microseta]; 2 eps, different in length; 1 long and 1 short ps; 1 long lsts; and 2 eus ( Figs. 3, 5, 6 View FIGURES 1 – 6 ). Abdominal segment IX with 3 ds, different in length; 1 very long ps; and 2 short sts, equal in length ( Figs. 4–6 View FIGURES 1 – 6 ). Lateral lobes of abdominal segment X with 1 minute ts.
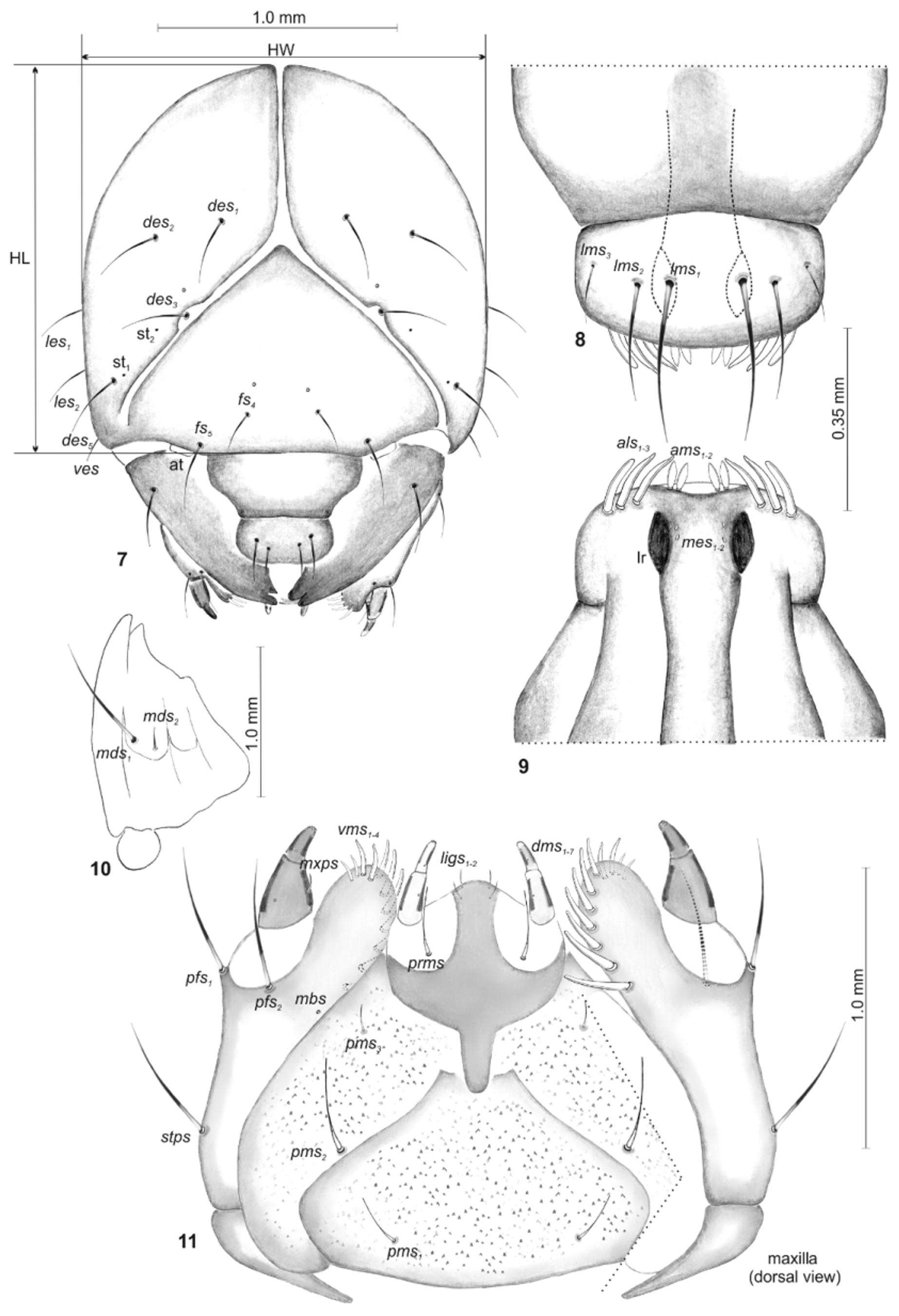
Head subglobose, slightly narrowed bilaterally ( Fig. 7 View FIGURES 7 – 11 ). Head capsule with 4 medium long des, des 1 and des 2 placed on central part of epicranium, des 3 located on frontal suture, des 5 located anterolaterally; 2 medium long fs, fs 4 placed anteromedially, fs 5 located near to antenna; 2 long les; and 1 short ves. Postepicranial area with a sensillum on each side, next pair of sensilla located medially on frons. Stemmata (2 pairs) feebly visible. Antennal segment membranous, bearing 1 conical sensorium and 3–4 filiform sensilla. Clypeus ( Fig. 8 View FIGURES 7 – 11 ) 2.7 times as wide as long, without setae. Labrum ( Fig. 8 View FIGURES 7 – 11 ) about 1.8 times as wide as long with 3 straight lms of different length, placed medially or mediolaterally; lms 3 distinctly shorter than other setae, lms 1 and lrm2 exceeding the outline of the labrum; the anterior margin of labrum almost rounded. Epipharynx ( Fig. 9 View FIGURES 7 – 11 ) with 3 finger-like als, different in length; 2 ams similar in both, shape and length, and 2 very short, poorly visible mes; labral rods (lr) short, reniform, almost parallel. Mandible ( Fig. 10 View FIGURES 7 – 11 ) bifid, teeth almost of equal height; with 2 mds, different in length; internal edge with a triangular tooth. Maxilla ( Fig. 11 View FIGURES 7 – 11 ) with 1 very long stps and 2 very long pfs, placed ventrolaterally; 1 very short mbs, situated ventrally. Mala with 7 dms, different in length and 4 straight vms, different in length. Maxillary palpi with two palpomeres; basal palpomere larger than distal; basal one with 1 mxps; distal palpomere with a group of 5 conical, cuticular apical processes; each palpomere with a sensillum. Praelabium heart-shaped ( Fig. 11 View FIGURES 7 – 11 ), with 2 very short ligs and 1 long prms. Labial palpi with two palpomeres, relatively elongated; basal palpomere slightly longer than distal; praemental sclerite well visible. Postlabium with 3 pms, different in length: pms 2 long, 3 times as long as pms 1, and 4 times as long as pms 3; surface of postlabium densely covered by distinct cuticular processes ( Fig. 11 View FIGURES 7 – 11 ).
Differential diagnosis. See “Key to larvae of selected Otiorhynchus species” and Tables 1, 2.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Entiminae |
|
Genus |