
Planothidium audax E.Morales, Novais & M.L.García, 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.510.3.9 |
|
persistent identifier |
https://treatment.plazi.org/id/03EA87A7-FF80-FFD6-FF2C-F920FABCFC6C |
|
treatment provided by |
Marcus |
|
scientific name |
Planothidium audax E.Morales, Novais & M.L.García |
| status |
sp. nov. |
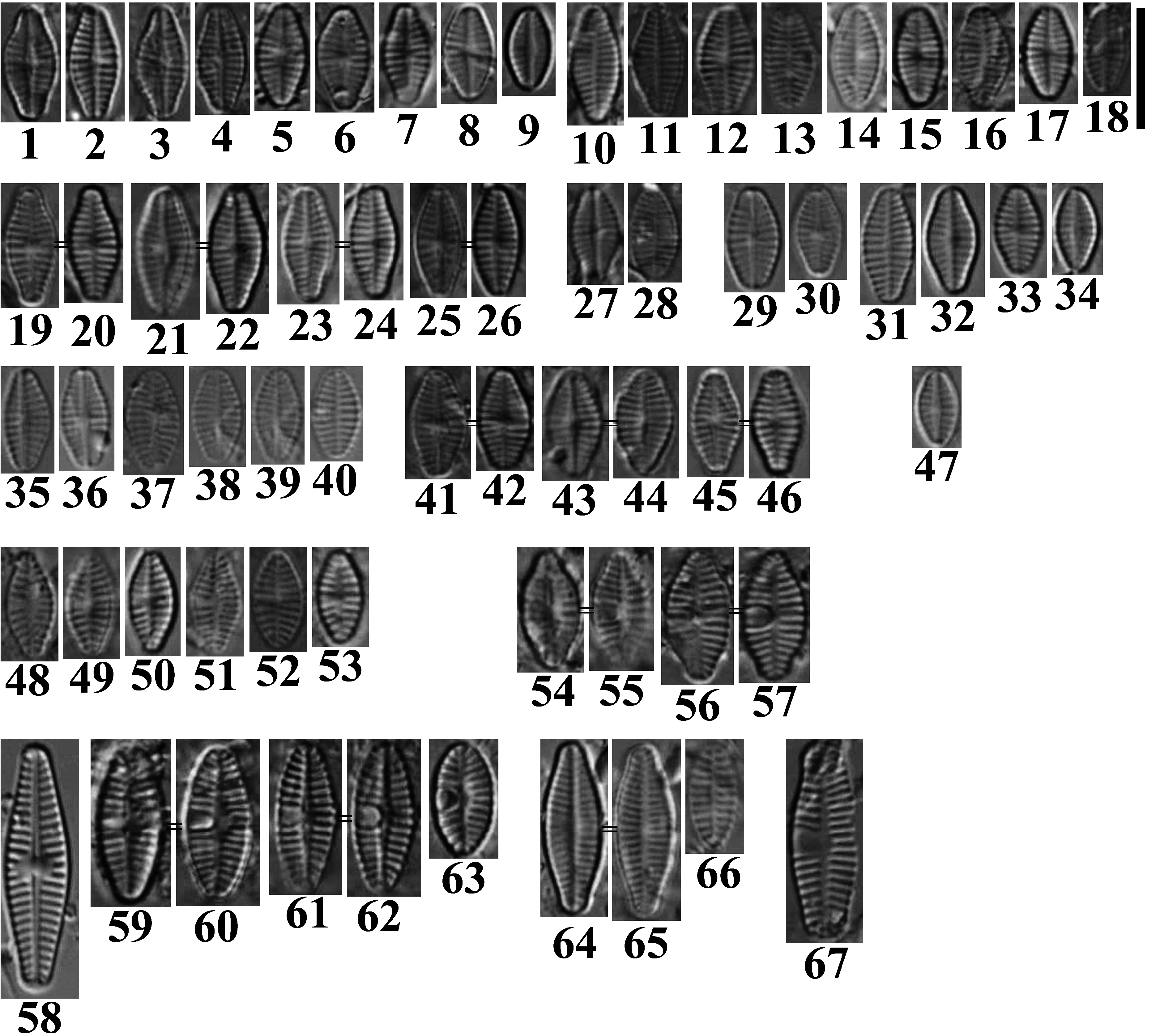
Planothidium audax E.Morales, Novais & M.L.García sp. nov. ( Figs 1–26 View FIGURES 1–67 LM, 68–73 SEM)
Valves lanceolate with convex margins and broadly rounded, rostrate ends. Smaller valves slightly elliptic lanceolate. Areolae round to elliptic and occluded by hymenes. Girdle elements open, non-perforated, very thin. Dimensions (n=30): length 7.1–10.1 µm, width 3.5–4.7 μm. Raphe valve (RV) ( Figs 1–9, 19, 21, 23, 25 View FIGURES 1–67 , 68, 69 View FIGURES 68–73 ): axial area linear to narrowly lanceolate, only slightly raised externally, widening at the central nodule where it connects to faintly squarish or rectangular central area, and clearly raised together with virgae in internal view. Central nodule elevated in internal view. Shortened striae frequently present on both sides of the central nodule, many times appearing as striae separated by a widened virga on primary valve side and two opposing widened virgae on the secondary side (best seen in Fig. 69 View FIGURES 68–73 ). Striae slightly radiate at central area to visibly radiate toward the valve ends; 18–20 in 10 μm. Striae commencing with 1–2 areolae at either valve face or mantle, becoming 3 rows only on valve face; running uninterruptedly onto valve mantle. Raphe filiform, slightly S-shaped, externally with straightly opposite proximal raphe ends and distal ends deflected in the same direction, barely extending onto valve mantle. Raphe internally with slightly deflected proximal ends in opposite directions. Helictoglossa faint, fused with apical valve thickening. Sternum valve (SV) ( Figs 10–18, 20, 22, 24, 26. 70–73 View FIGURES 1–67 View FIGURES 68–73 ): axial area linear to narrowly lanceolate, flat in external view, raised in internal view and at the same level as virgae. Striae of same characteristics as those on RV, shortened striae may or may not be present on both sides of the central nodule; 17–22 in 10 μm.
Type:— PORTUGAL. Alentejo region , Serpa Council: Limas Stream, sampling point in the stream, down from main road bridge, 37º 49’ 19.91” N, 7º 37’ 21.18” W, M.H. Novais, 20th September 2017 (holotype BR-4663 = Figs. 21 and 22 View FIGURES 1–67 , same frustule) GoogleMaps .
Etymology:—The epithet “ audax ” refers to the ability (audacity) of the species to live in the extreme conditions of the
Limas Stream, a temporary ecosystem.
Ecology and distribution:— The new species was present (1% in a count of 600 valves) in a sample collected by scrubbing 5 hand-sized rocks from a pool within the dry riverbed during summer, the dry season in southern Portugal (for collection and analysis methods see Novais et al. 2020) ( Figs 1–26 View FIGURES 1–67 ). The stream bed was mostly unconsolidated with very large, spaced boulders, 60% dry, no shading, and presented some filamentous algal growth in zones with water. The sampled pool was 5× 2 m in size, 0.3 m deep, and the turbidity was 7.6 NTU. Water temperature was 33.8 ºC (at 12:00 am), pH 8.8, electrical conductivity 752.5 µS∙cm, dissolved oxygen 79.8% sat., phosphates 0.2 mg PO 4 ∙L- 1, and nitrates 1.9 mg NO 3 ∙L- 1 .
-1
Major impacts on the stream (besides the seasonality of precipitation being higher during winter and causing a high degree of erosion and sediment transport, filling the ca. 10 m-wide stream bed from side to side and ca. 2 m above the bottom) are a road and a bridge that cross the stream just below the sampling point, and cattle raising.
In samples collected on the same date, the new species was found in a nearby pool reaching an abundance of 0.5% and also in a sample of dry biofilm (in this case the taxon was not recorded during the count). The dry biofilm was collected scrubbing the covered surface of the same number of rocks as the pool samples, but taken randomly from the dry riverbed. The second pool had a water temperature of 27.6 ºC (at 12:00 am), pH 8.7, electrical conductivity 946.0 µS∙ cm-1, dissolved oxygen 68.5% sat., phosphates 0.1 mg PO 4 ∙L- 1, and nitrates 2.7 mg NO 3 ∙L- 1. No chemistry data is available for the dry biofilm sample.
The new species was found in highest abundance (2.8%) in a pool sample from the Maria Delgada Stream (Serpa Council, 37º 39’ 36,42” N, 7º 37’ 21,18” W, collected 19 th September 2017, Figs 41–46 View FIGURES 1–67 ), but it had low quality for SEM analysis. In this case, the pool was 0.5 m deep, temperature 15 ºC (at ca. 10 am), pH 8.9, electrical conductivity 2280.0 µS∙ cm-1, dissolved oxygen 45.7% sat., phosphates 0.1 mg PO 4 ∙L- 1 and nitrates 9.4 mg NO 3 ∙L- 1 GoogleMaps .
In order to provide a sense of temporal and spatial variability of P. audax sp. nov., material from several dates collected from the type locality and from the Maria Delgada Stream are provided ( Figs 1–47 View FIGURES 1–67 ). In the latter figures it can be observed that the morphological variability of the taxon tends to remain relatively stable in the different dates and in the two streams (for collection dates and stream information see legend to Figs. 1–67 View FIGURES 1–67 ), with the exception that in a pool sample from the Maria Delgada Stream, we found only one valve, but the smallest RV (5.9 × 3.1 µm and 17 striae per 10 µm, Fig. 47 View FIGURES 1–67 ) in 28 th May 2018 .
Accompanying flora:—In the type sample the new species was initially identified as Planothidium cf. granum . The most abundant accompanying species were Pseudostaurosira sp. 1 (36.9%), Pseudostaurosiropsis sp. 2 (11.7%), Pseudostaurosira sp. 3 (6.8 %), Amphora pediculus (Kützing) Grunow (in Schmidt 1875: pl. 26, fig. 99, 4.9%), Stauroforma atomus (Hustedt) D.Talgatti, C.E.Wetzel, E.Morales & L.C.Torgan (in Talgatti et al. 2014: 45, 4.4%), Epithemia adnata (Kützing) Brébisson (1838:16 , 4.2%), Epithemia sorex Kützing (1844: 33 , 3.7%) and other species present in abundances <2.9%. Undetermined species are currently under description.
Regarding other Planothidium species in the type sample, we found P. delicatulum (Kützing) Round & Bukhtiyarova (1996: 353 , not pictured since only broken valves were found), P. engelbrechtii (Cholnoky) Round & Bukhtiyarova (1996: 353 , Figs 64–66 View FIGURES 1–67 ), P. lanceolatum (Brébisson ex Kützing) Lange-Bertalot (1999: 287 , Fig. 67 View FIGURES 1–67 ), P. minutissimum (Krasske) E. Morales (2006: 338 , Figs 48–53 View FIGURES 1–67 ), P. rostratoholarcticum Lange-Bertalot & Bąk in Bąk & Lange-Bertalot (2014: 354, Figs 54–57 View FIGURES 1–67 ), and P. victorii Novis, Braidwood & Kilroy (2012: 22 , Figs 58–63 View FIGURES 1–67 ). Of these, only the latter reached an abundance of 1.9%, while the remaining taxa were not recorded during the count.
In the Maria Delgada Stream pool sample in which the taxon was more abundant, the accompanying flora was represented by Pseudostaurosira sp. 3 (25.9%), P. sp. 1 (11.9%), Amphora pediculus (Kützing) Grunow in A.W.F. Schmidt (1875: pl. 26, fig. 99, 7.4%), Pseudostaurosiropsis sp. 2 (7.2%), Nitzschia valdecostata Lange-Bertalot & Simonsen (1978: 58 , 6.2%), Pseudostaurosiropsis sp. 1 (5.4%), Karayevia clevei (Grunow) Bukhtiyarova (1999: 94 , 5.4%), Cocconeis neothumensis Krammer (1990: 151 , 3.6%), Nitzschia inconspicua Grunow (1862: 579 , 3.2%) and other taxa in abundances of 2.4% or less.
Comments:—Following is a comparison with morphologically closely related species currently ascribed to Planothidium ; only differences are annotated.
In P. daui (Foged) Lange-Bertalot (1999: 275) larger valves are longer and wider (length 7–13, width 3.6–5.8) and striation is coarser on both valves, 14–17 in 10 μm. Valves are lanceolate with subcapitate to capitate ends. In the RV, the axial area is internally and externally raised, it has a strongly radiate striation pattern, with clear bow tie central area. Externally, the distal raphe ends are deflected in the same direction and continue deep into the mantle. The helictoglossa is fused with an apical valve thickening, but it is raised above it. The striae have up to 4 rows of areolae and only slightly extend onto the mantle. In the SV the axial area and virgae are externally depressed, and the striae are composed of up to 6 rows of areolae.
In P. engelbrechtii larger valves become longer and wider (length: 6.2–20, width 3.3–5.2, Compère & Van de Vijver 2009), and the RV can have coarser striae (16.5–20 in 10 µm). The RV has an axial area and virgae raised externally and the raphe distal ends are deflected in the same direction, running deep into the valve mantle. Internally, the raphe is S-shaped with tilted helictoglossa. The striae are restricted to the valve face. The SV has an internally clearly depressed axial area and very thin elevated costae. The striae have up to 4 rows of areolae.
In P. granum (Hohn & Hellerman) Lange-Bertalot (1999: 272) valves are lanceolate with accentuated rostrate ends and the striation is coarser with 15–17 in 10 µm on both valves. The RV has an accentuated radiate striation pattern, a clear bow tie central area, and striae are mostly on the valve face and only one or two areolae are on the valve mantle. The helictoglossa is fused with an apical thickening, but it is raised above it. Internally, the proximal raphe ends are hooked and clearly deflected in opposite directions. The SV is externally depressed together with the virgae, and the striae have up to 7 rows of areolae.
In P. minutissimum the valves are more lanceolate to rhombic with somewhat cuneate or broadly rounded ends. Striation is coarser on both RV and SV, being 14 to17 and 14–16 in 10 µm, respectively ( Morales 2006, Stancheva et al. 2020). The striation pattern in the RV is much more radiate with a wider central area. Externally, the raphe is strongly deflected in a hook-like pattern at both valve ends and it penetrates deeply into the valve mantle. The striae stop at valve face/mantle junction or continue onto it only with a few areolae. The helictoglossa are higher and do not associate with the apical valve thickening. The SV has an axial area sunken in external view, but raised and higher that virgae in internal view. There is a clear spacing (separation of striae) on one side of central area. Striae has same pattern as in RV.
In P. pumilum Bąk & Lange-Bertalot (2014: 350) the shape of valves tends to be more sublanceolate, elliptical in many cases, with subrostrate apices. The striation in RV and SV is coarser, 15 in 10 μm and 15.5–16.5 in 10 μm, respectively. The RV valve has a wider, rectangular central area. Externally, the raphe goes deeper into mantle at apices and internally, the helictoglossa is independent from the apical thickening. The striae stop at the valve face/mantle junction and are formed by 3–4 rows of areolae. The SV has recessed axial area and virgae in outer view. The striae are slightly parallel in the middle to radiate at the end, and are formed by up to 4 rows of areolae.
In P. rhombiculum Lange-Bertalot, Bąk & Hofmann (in Bąk & Lange-Bertalot 2014: 353), valves have a wider axial area. The RV and SV have coarser striae, being 12.5–13.5 in 10 μm in the central part and 14–15.5 in 10 μm near the ends in the RV and 15–18 (measured from SEM images in Bąk & Lange-Bertalot 2014) in the SV valve. In the RV, the raphe has externally a clear S-shape with proximal raphe ends deflected in the same direction. The striae are pointier at the axial area because they tend to be uniseriate as they approach it. The helictoglossa are tilted and fused with a less prominent apical thickening. In the SV valve the axial area is recessed in outer view.
From the comparison above we deduct that the stria density on the SV (17–22 in 10 μm), combined with the lanceolate valves (becoming elliptic lanceolate in smaller individuals), with convex margins and broadly rounded, rostrate ends are distinguishing characters of P. audax sp. nov. It must be also mentioned that, although the following are features seldom mentioned for monoraphid diatoms, the new taxon has a unique combination in the flat external configuration of the complex axial area-virgae-striae in outer view, and the internally raised axial area-virgae with respect to the recessed stria; this configuration occurs in RV and SV. Because of our finding of the smallest RV in a sample other than the type, the size range for this valve is expanded to length 5.9–10.1 µm, width 3.1–4.7 μm, while the stria density becomes 17–20 in 10 μm .

| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Planothidium audax E.Morales, Novais & M.L.García
| Morales, Eduardo A., Novais, Maria Helena, García, María Luján, Maidana, Nora I. & Morais, Maria Manuela 2021 |
P. pumilum Bąk & Lange-Bertalot (2014: 350)
| Bak, M. & Lange-Bertalot, H. 2014: ) |
P. daui (Foged)
| Lange-Bertalot, H. 1999: ) |
P. granum (Hohn & Hellerman)
| Lange-Bertalot, H. 1999: ) |