
Phaeophilacris (Taenacris) kereita, Carvalho & Castro-Souza & Njoroge & Kinuthia & Ferreira, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5433.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:149299C6-B272-46D1-8389-46342C2CAA6C |
|
DOI |
https://doi.org/10.5281/zenodo.10954407 |
|
persistent identifier |
https://treatment.plazi.org/id/03E98790-495C-FFCD-FF68-F8F0FBB5FA9A |
|
treatment provided by |
Plazi |
|
scientific name |
Phaeophilacris (Taenacris) kereita |
| status |
sp. nov. |
Phaeophilacris (Taenacris) kereita n. sp.
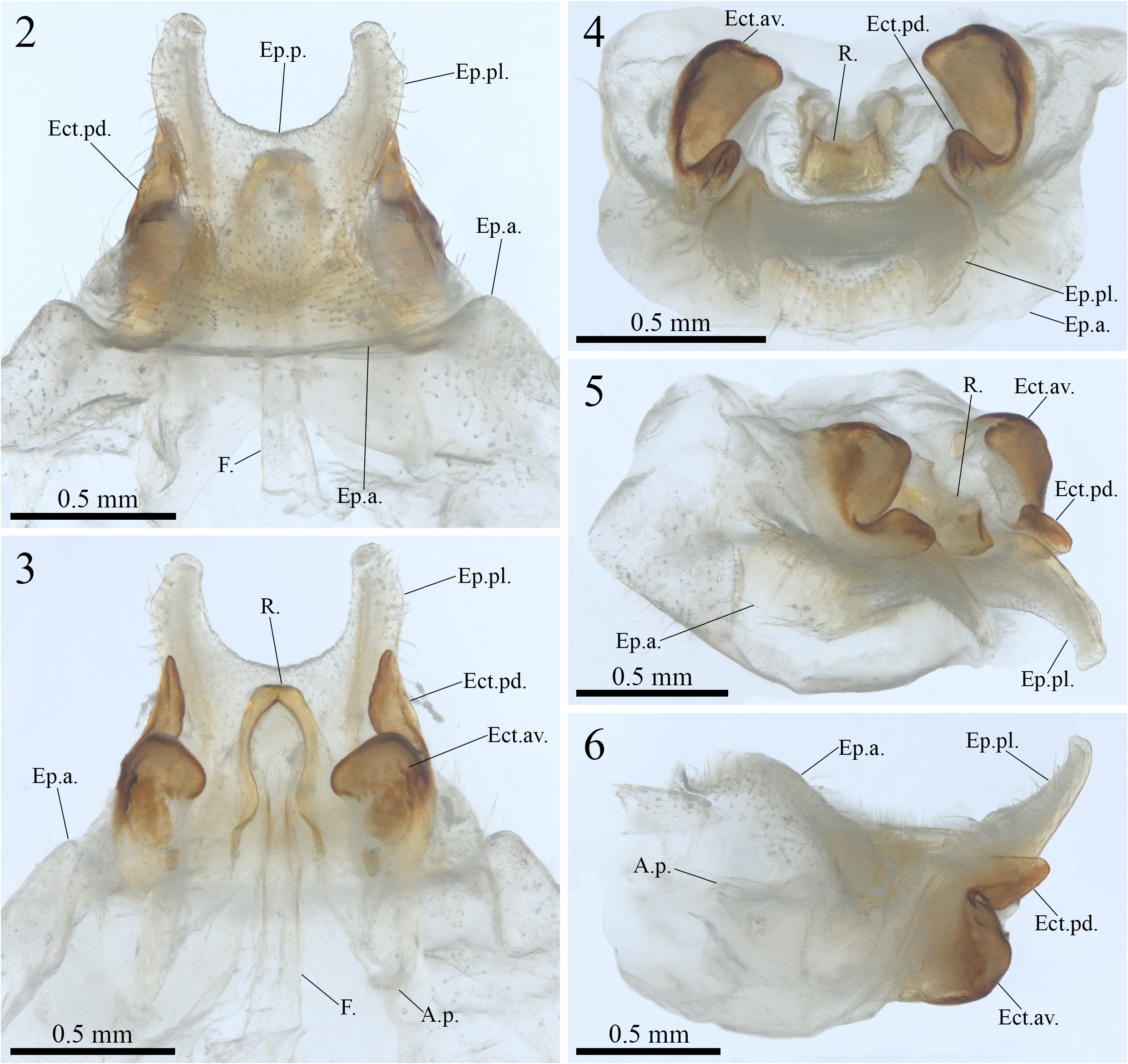
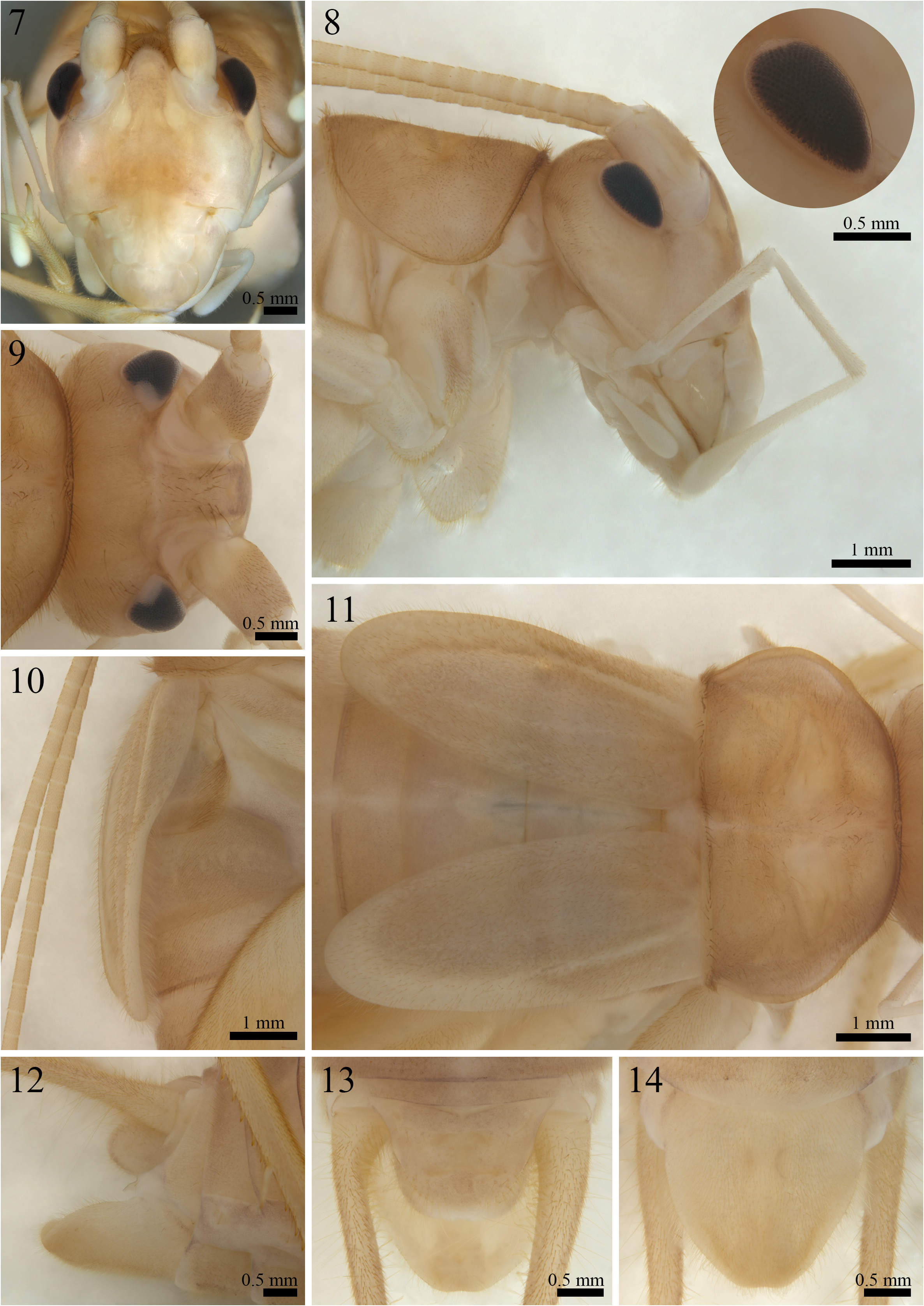
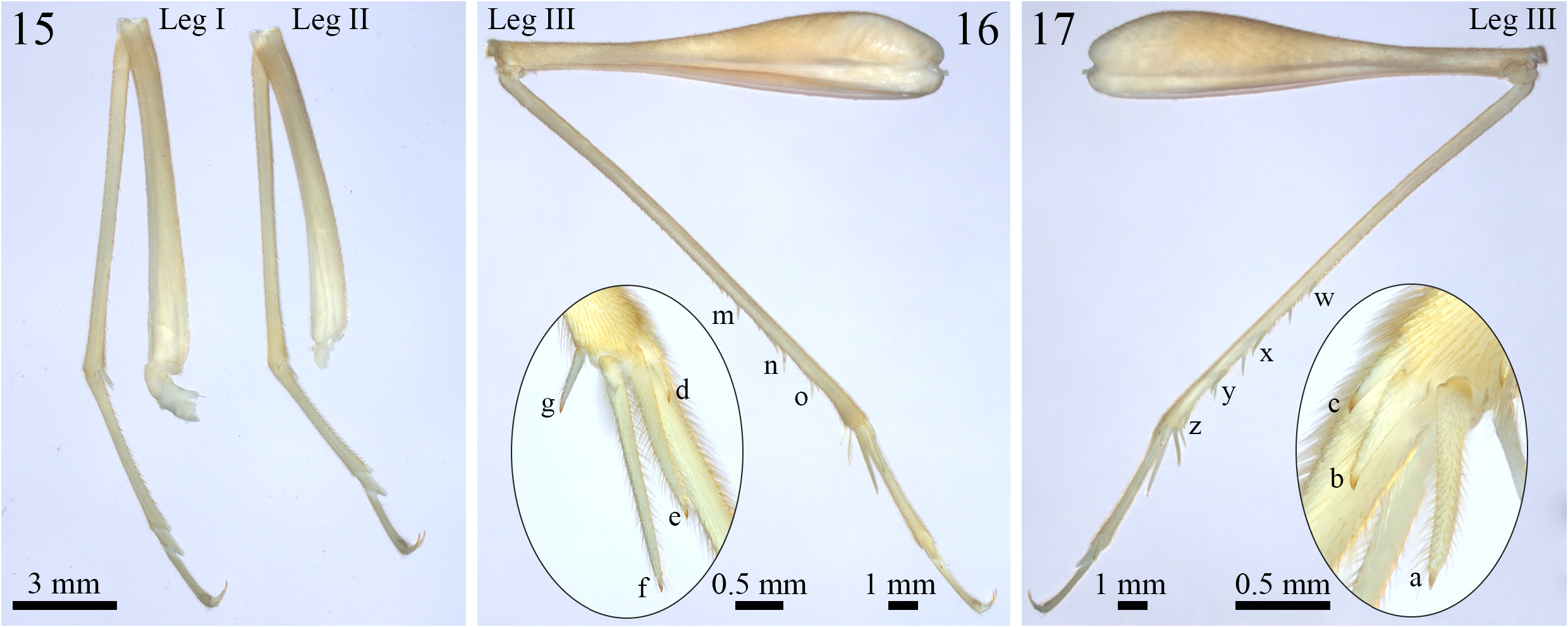
( Figures 2–6 View FIGURES 2–6 , 7–14 View FIGURES 7–14 , 15–17 View FIGURES 15–17 , 18–22 View FIGURES 18–22 ; Table 1 View TABLE 1 )
Material examined— Holotype ( NMK/INV/T-748 ), ♂, Kenya, Gatamaiyu Forest Natural Reserve , Kereita cave (36°39’46.62”E, 0°56’3.03”S), 24.I.2020, R.L. Ferreira; condition: body coloration tarnished, male genitalia dissected and stored alongside the holotype, left legs I, II and III, and right leg III detached and also stored alongside the holotype GoogleMaps . Paratype ( NMK/INV/T-749 ), ♂, Kenya, Gatamaiyu Forest Natural Reserve , Kereita cave (36°39’46.62”E, 0°56’3.03”S), 24.I.2020, R.L. Ferreira; condition: body coloration slightly tarnished, male genitalia dissected and stored alongside the holotype, left legs I, II and III detached and also stored alongside the paratype, right leg III missing GoogleMaps .
Diagnosis— Combination of the following characteristics: posterior portion of the epiphallus membranous and U-shaped ( Figs. 2 and 3 View FIGURES 2–6 , Ep.p.); posterolateral lobes of the epiphallus short and dorsally projected, apex subtriangular and also dorsally projected ( Figs. 2, 5 and 6 View FIGURES 2–6 , Ep.pl..); ectoparameres divided into anteroventral and posterodorsal lobes, posterior margin of the anteroventral lobe with a short, centralized and rounded projection pointing towards the posterior region of the genitalia, posterodorsal lobe tapered, slightly curving away from the center of the genitalia and roughly V-shaped ( Figs. 3, 5 and 6 View FIGURES 2–6 , Ect.av. and Ect.pd.); rachis with an anteromedial constriction, forming a ring around the posterior portion of the formula, posterior margin quadrangular ( Fig. 3 View FIGURES 2–6 , R.).
Male genitalia (holotype, Figs. 2–6 View FIGURES 2–6 )—phallic complex short in general view ( Figs. 2–6 View FIGURES 2–6 ). Epiphallus: anterior portion dorsally projected and inclined towards the anterior region of the body, lateral margins rounded and expanding, to a small extent, towards the posterior end of the genitalia ( Figs. 2 and 6 View FIGURES 2–6 , Ep.a.); posterior portion membranous and U-shaped ( Figs. 2 and 3 View FIGURES 2–6 , Ep.p.); posterolateral lobes short, predominantly membranous, with a slightly sclerotized band extending along their entire length, dorsally projected (in lateral view) and curved towards the center of the genitalia (in dorsal and ventral views), apex subtriangular and also dorsally projected, with dorsolateral surface covered with setae, lateral setae elongated ( Figs. 2, 3 and 6 View FIGURES 2–6 , Ep.pl.); rami missing (Rm.). Ectoparameres: sclerotized and divided into anteroventral lobe and posterodorsal lobe ( Figs. 3–6 View FIGURES 2–6 , Ect.av. and Ect.pd.); anteroventral lobe well-developed, expanding both ventrally and laterally, with an almost quadrangular contour in lateral view, inner margin with a medium-sized rounded projection, posterior margin with a short, centralized, and rounded projection pointing towards the posterior region of the genitalia ( Figs. 3–6 View FIGURES 2–6 , Ect.av.); posterodorsal lobe tapered, extending posteriorly, reaching in length the posterior margin of the epiphallus and slightly curving away from the center of the genitalia, with a shape similar to that of a fin in ventral view, outline roughly V-shaped in lateral and laterofrontal views ( Figs. 3–6 View FIGURES 2–6 , Ect.pd.). Endoparamere: apodema principale membranous, concave (in lateral view), extremities slightly dorsally projected ( Fig. 3 View FIGURES 2–6 , A.p.). Rachis: almost as long as the central margin of the posterior portion of the epiphallus, sclerotized, with an anteromedial constriction, forming a ring around the posterior portion of the formula, posterior margin with a quadrangular outline (in ventral view) and small acute centered projection in frontal view ( Figs. 3 and 4 View FIGURES 2–6 , R.). Formula: almost as long as the apodema principale, rectangular-shaped and membranous, region adjacent to the constriction of the rachis smaller in diameter and slightly sclerotized ( Fig. 3 View FIGURES 2–6 , F.).
Variations in the male genitalia (paratype)— Epiphallus: posterolateral lobes (Ep.pl.) with a sclerotized and wide band extending along their entire length, dorsally projected but not curved towards the center of the genitalia; rami (Rm.) slightly sclerotized, elongated, longer than the apodema principale. Endoparamere: apodema principale (A.p.) slightly sclerotized. Formula (F.) shorter than the apodema principale, rectangular and sclerotized, with the margin adjacent to the constriction of the rachis being more sclerotized than the rest of the structure.
Body morphology (holotype, Figs. 7–17 View FIGURES 7–14 View FIGURES 15–17 )— Body color (after fixation in ethanol 70%): head, pronotum, tegmina, abdomen, cerci and legs pale whitish yellow, without dark spots ( Figs. 7–14 View FIGURES 7–14 ); Body color (in vivo observations): general coloration pale brown, locomotor appendages ranging from pale brown to yellowish-orange ( Figs. 21 and 22 View FIGURES 18–22 ). Head: slightly pubescent, dorsoventrally elongated ( Figs. 7 and 8 View FIGURES 7–14 ), fastigium covered by long setae and approximately as wide as half the length of the antennal scape ( Fig. 9 View FIGURES 7–14 ); scape well-developed, 1.5 times longer than the compound eyes in lateral view ( Fig. 8 View FIGURES 7–14 ); maxillary palp divided into five palpomeres, the first two short and of similar length, palpomeres III and IV elongated and of comparable length, the terminal palpomere longer than the others with a club-shaped apex ( Fig. 8 View FIGURES 7–14 ); labial palp short and divided into three parts, the posterior palpomere as long as the anterior and median palpomeres combined, featuring a rounded and dilated apex ( Fig. 8 View FIGURES 7–14 ); ocelli absent, and the compound eyes slightly reduced, dorsal region near the antennal cavities depigmented ( Figs. 7–9 View FIGURES 7–14 ). Thorax: pronotum pubescent, with its anterior and posterior margins arched and covered by elongated bristles, the median region intersected by an anteroposterior groove, lateral lobes subtriangular, facing downward and slightly towards the head ( Figs. 8 and 11 View FIGURES 7–14 ); metanotal gland poorly developed, confined to the anterior margin of the metanotum, adorned with numerous setae, concealed by the tegmina, and subdivided into left and right lobes, which are separated by a whitish line that extends anterodorsally until reaching the supra-anal plate ( Fig. 13 View FIGURES 7–14 ). Abdomen: tergites pubescent; supra-anal plate subtriangular and pubescent, with a rounded posterior margin covered by long bristles, shorter than the subgenital plate and equal in length to the paraprocts ( Figs. 12 and 13 View FIGURES 7–14 ); subgenital plate approximately two times longer than the supra-anal plate, pubescent, with lateral and posterior margins also covered by long bristles, subtriangular-shaped, apex of the posterior margin straight ( Figs. 12 View FIGURES 7–14 to 14); cerci pubescent and covered by long setae throughout their length, the inner portion of the base covered by globose setae ( Figs. 12 View FIGURES 7–14 to 14). Tegmina: slightly sclerotized and moderately translucent, pubescent, reduced, and lanceolate, rounded at the apex and partially covering tergites I and II, as well as the anterior portion of tergite III; inner margins not touching; dorsal field intersected by two or three poorly-marked longitudinal veins, while the lateral field is bordered by an also poorly-marked longitudinal vein curving towards the center of the body, posterior portion of the lateral field whitish and thickened ( Figs. 10 and 11 View FIGURES 7–14 ). Legs: pubescent; leg I (28.01 mm) longer than leg II (25.55 mm), femur bearing small forward-pointing denticles on both ventral margins, tibia devoid of tympanum ( Fig. 15 View FIGURES 15–17 ); tibia of legs I and II bearing two ventral apical spurs ( Fig. 15 View FIGURES 15–17 ); tarsomere I of legs I (4.92 mm) and II (3.94 mm) serrated ventrally and longer than tarsomeres II and III combined ( Fig. 15 View FIGURES 15–17 ); leg III (41 mm) longer than legs I and II, tibia serrated dorsally, bearing four subapical spines on the outer dorsal margin ( Fig. 17-w, x, y, z View FIGURES 15–17 ) and three on the inner dorsal margin ( Fig. 16-m, n, o View FIGURES 15–17 ), as well as three apical spurs on the outer surface ( Fig. 17-a, b, c View FIGURES 15–17 ) and four on the inner surface ( Fig. 16-d, e, f, g View FIGURES 15–17 ); spur “a” longer than the others, spur “b” longer than spur “c”; spur “f” longer than the others, spur “e” longer than spurs “d” and “g”, tarsomere I (4.61 mm) of leg III also serrated ventrally, longer than tarsomeres II and III together, and bearing two small dorsal subapical spines and two apical spurs, with the inner one longer than the outer one ( Figs. 16 and 17 View FIGURES 15–17 ).
Variations in body morphology (paratype)— Legs: tibia of left leg III bearing five subapical spines instead of four on the outer margin.
Ecological remarks— Kereita cave is located within the limits of the Gatamaiyu Forest Natural Reserve. Gatamaiyu Forest ( Fig. 18 View FIGURES 18–22 ) is a remaining section of the Kikuyu Escarpment Forest, encompassing a vast area of 37,620 hectares. This expansive forested landscape comprises several distinct forest blocks, including Kereita, Kinale, Kamae, Kieni, Raggia, Kijabe, and Uplands, in addition to Gatamaiyu. Within Gatamaiyu Forest, which spans 4,720 hectares, approximately 75% consists of native forest, while 8% is characterized by exotic tree species. The remaining portion features scrublands, bamboo groves, and herbaceous croplands. Despite extensive logging activities occurring in the 1950s and 1960s, Gatamaiyu Forest has remarkably regenerated over time. This forest serves a vital role as a watershed area for numerous rivers and springs, including the Gatamaiyu, Bathi, Kiruiru, and Nyanduma rivers.
The entrance to Kereita Cave is situated at an altitude of 2,400 meters above sea level, nestled in the heart of the forest, adjacent to a substantial waterfall ( Fig. 19 View FIGURES 18–22 ). Due to its proximity to the lake created by the waterfall, it is likely that during periods of heavy rainfall, the lower main chamber of the cave may experience flooding. Nevertheless, the cave floor in the main chamber is consistently damp due to the mist generated by the waterfall, which permeates through the cave entrance.
The cave consists of a main chamber, approximately 20 meters in length, and a secondary upper conduit, spanning approximately 10 meters. Notably, individuals of the species Phaeophilacris (T.) kereita n. sp. were not observed within the lower main cave chamber. Instead, they were primarily found in the lateral upper conduit, particularly along the cave’s ceiling and walls. These specimens tended to cluster around cracks in the rock formations ( Fig. 21 View FIGURES 18–22 ). It is worth mentioning that the majority of the specimens observed in these aggregations were immatures, with only two adult males identified ( Fig. 22 View FIGURES 18–22 ), despite the presence of several sub-adult male specimens.
The primary organic materials present in the cave included plant debris carried in by water and bat guano. An intriguing aspect of the cave environment is the intense noise level caused by the nearby waterfall. The noise is so pronounced that even holding conversations inside the cave becomes challenging.Consequently, any form of acoustic communication by the crickets is impeded. The behavior of these specimens was particularly intriguing. Typically, cave crickets tend to exhibit evasive behavior, scurrying away when approached with headlamps, suggesting a degree of photophobia and a tendency to seek shelter when disturbed. However, what sets Phaeophilacris (T.) kereita n. sp. apart is the surprising composure of these specimens: they remained remarkably calm, even in the presence of a human with powerful lights in close proximity. Unlike other species, they did not display any attempts to seek shelter but rather remained still and composed.
Although the waterfall near Kereita Cave is a tourist attraction, the cave itself appears to receive very few visitors. To access the cave, one must cross the river, as its entrance is on the opposite bank from the trail. Additionally, because the cave entrance is in close proximity to the waterfall, many visitors seem hesitant to enter, often choosing to avoid it. As a result, the interior of the cave remains relatively undisturbed, showing minimal signs of human interference. Furthermore, access to the cave is only permitted when visitors are accompanied by park rangers, as the area is known to have elephants and other wildlife that could pose a risk to humans.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |