
Phaeophilacris (Taenacris) papiophagus, Carvalho & Castro-Souza & Njoroge & Kinuthia & Ferreira, 2024
|
publication ID |
https://doi.org/10.11646/zootaxa.5433.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:149299C6-B272-46D1-8389-46342C2CAA6C |
|
DOI |
https://doi.org/10.5281/zenodo.10954414 |
|
persistent identifier |
https://treatment.plazi.org/id/03E98790-4956-FFCB-FF68-FA5DFAE1FEA2 |
|
treatment provided by |
Plazi |
|
scientific name |
Phaeophilacris (Taenacris) papiophagus |
| status |
sp. nov. |
Phaeophilacris (Taenacris) papiophagus n. sp.
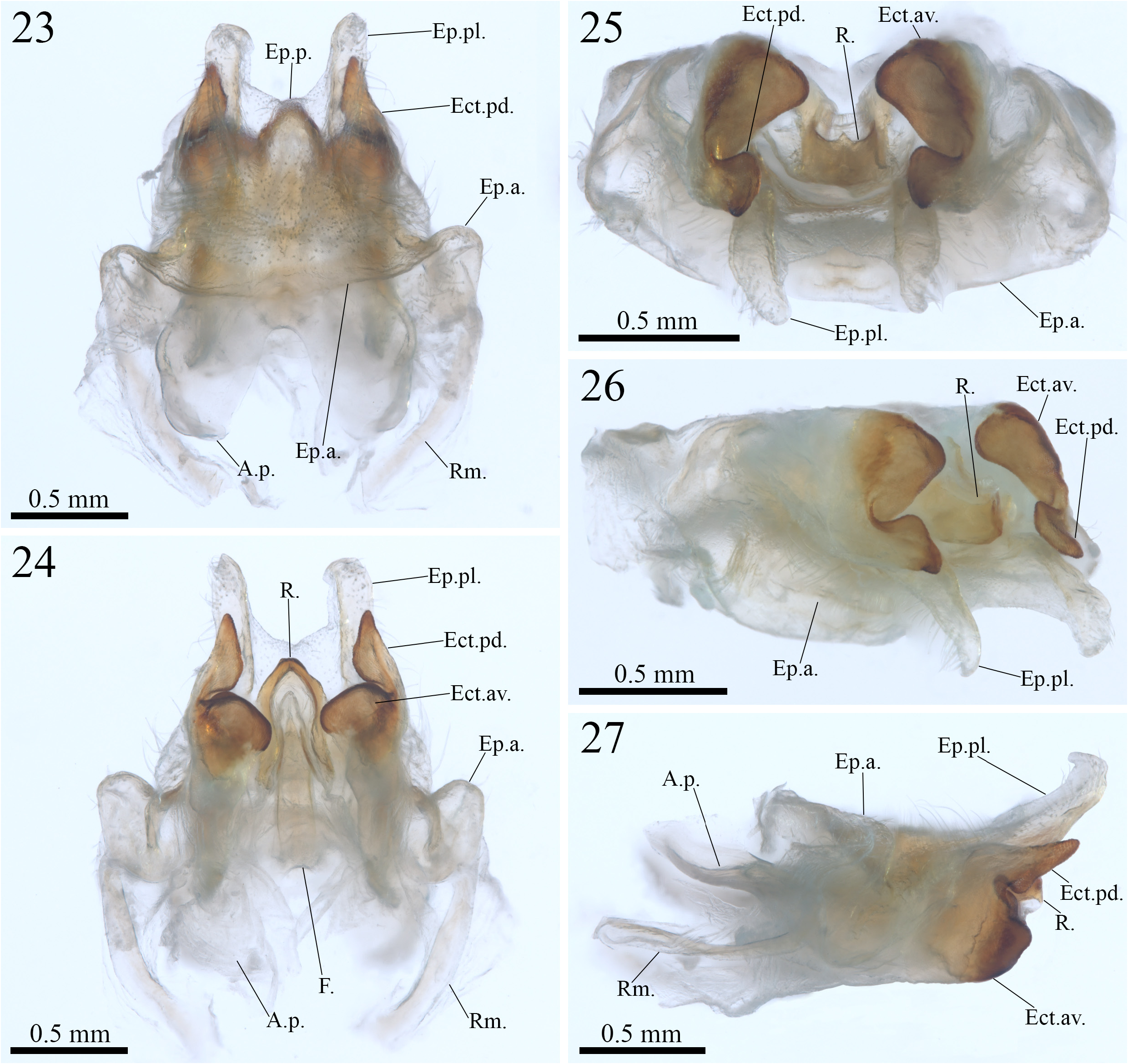
( Figures 23–27 View FIGURES 23–27 , 28–32 View FIGURES 28–32 , 33–36 View FIGURES 33–36 ; Table 2 View TABLE 2 )
Material examined— Holotype ( NMK/INV/T-750 ), ♂, Kenya, Rift Valley , Mount Suswa, Baboons cave (36°24’17.168”E, 1°7’54.422”S), 23.I.2020, R.L. Ferreira; condition: body coloration slightly tarnished, male genitalia dissected and stored alongside the holotype, subgenital plate, left legs I and II detached and also stored alongside the holotype, left leg III, and right legs I and III missing. GoogleMaps
Diagnosis— Combination of the following characteristics: posterior portion of the epiphallus membranous and slightly U-shaped ( Fig. 24 View FIGURES 23–27 , E.p.); posterolateral lobe of the epiphallus short and dorsally projected, apex subtriangular and also dorsally projected, tip facing the anterior region of the phallic complex ( Figs. 24, 26 and 27 View FIGURES 23–27 , Ep.pl.); ectoparameres divided into anteroventral and posterodorsal lobes, posterior margin of the anteroventral lobe with a short, centralized, rounded and more pronounced projection pointing towards the posterior region of the genitalia, posterodorsal lobes more tapered than those of P. (T.) kereita n. sp., ventral margin with a small, but pronounced rounded projection adjacent to the posterior margin of the anteroventral lobe ( Figs. 24–27 View FIGURES 23–27 , Ect.av. and Ect.pd.); rachis with an anteromedial constriction, forming a ring around the posterior portion of the formula, posterior margin subtriangular ( Figs. 24, 25 and 27 View FIGURES 23–27 , R.).
Male genitalia (holotype, Figs. 23–27 View FIGURES 23–27 )—phallic complex short in general view ( Figs. 23–27 View FIGURES 23–27 ). Epiphallus: anterior portion minimally dorsally projected and highly inclined towards the anterior region of the body in comparison to P. (T.) kereita n. sp., lateral margins rounded and expanding, to a small extent, towards the posterior end of the genitalia, akin to P. (T.) kereita n. sp. ( Figs. 23 and 27 View FIGURES 23–27 , Ep. a.); posterior portion membranous and Ushaped to a lesser extent when compared to that of P. (T.) kereita n. sp. ( Figs. 23 and 24 View FIGURES 23–27 , Ep. p.); posterolateral lobes shorter than those of P. (T.) kereita n. sp., but not curved inwards (in dorsal and ventral views), predominantly membranous, with a sligthly sclerotized band extending along their entire length, apex subtriangular and dorsally projected, similar to P. (T.) kereita n. sp., but with the tip facing the anterior region of the phallic complex, with the dorsolateral surface covered with setae, lateral setae elongated, similar to P. (T.) kereita n. sp. ( Figs. 23, 24, 26 and 27 View FIGURES 23–27 , Ep. pl.); rami present, slightly sclerotized, elongated, longer than the apodema principale ( Figs. 23, 24 and 27 View FIGURES 23–27 , Rm.). Ectoparameres: sclerotized and divided into anteroventral lobe and posterodorsal lobe ( Figs. 24–27 View FIGURES 23–27 , Ect.av. and Ect. pd.); anteroventral lobe developed, expanding both ventrally and laterally, with an almost quadrangular contour that is more defined than that of P. (T.) kereita n. sp., rounded projections situated in the inner and posterior margins are also more pronounced than those of P. (T.) kereita n. sp. ( Figs. 24–27 View FIGURES 23–27 , Ect.av.); posterodorsal lobe more tapered than that of P. (T.) kereita n. sp., extending towards the posterior region of the genitalia, slightly surpassing the posterior margin of the epiphallus in length and slightly curving away from the center of the genitalia, with a shape similar to that of a fin in ventral view, outline roughly V-shaped in lateral and laterofrontal views, ventral margin with a small, but pronounced rounded projection adjacent to the posterior margin of the anteroventral lobe ( Figs. 24–27 View FIGURES 23–27 , Ect. pd.). Endoparamere: apodema principale slightly sclerotized, concave (in lateral view), extremities slightly dorsally projected ( Figs. 23 and 27 View FIGURES 23–27 , A.p.). Rachis: almost as long as the central margin of the posterior portion of the epiphallus, sclerotized, with an anteromedial constriction, forming a ring around the posterior portion of the formula, resembling P. (T.) kereita n. sp., posterior margin subtriangular, with a more tapered outline and small acute centered projection in frontal view ( Figs. 24 and 25 View FIGURES 23–27 , R.). Formula: shorter than the apodema principale, rectangular in shape and sclerotized, with the region adjacent to the constriction of the rachis being more sclerotized and smaller in diameter than the rest of the structure ( Fig. 24 View FIGURES 23–27 , F.).
Body morphology (holotype, Figs. 28–32 View FIGURES 28–32 )— Body color (after fixation in ethanol 70%): head, pronotum and tegmina ranging from yellowish-orange to brown ( Figs. 28–30 and 32 View FIGURES 28–32 ), abdomen, cerci and legs (I and II) pale whitish-yellow; Body color (in vivo observations): general coloration dark brown, locomotor appendages light yellowish-red. Head: slightly pubescent, dorsoventrally elongated ( Figs. 28 and 29 View FIGURES 28–32 ), fastigium covered by long setae and approximately as wide as half the length of the antennal scape ( Fig. 28 View FIGURES 28–32 ); scape well-developed, 1.5 times longer than the compound eyes in lateral view ( Fig. 29 View FIGURES 28–32 ); maxillary palp divided into five palpomeres, the first two short and of similar length, palpomeres III and IV elongated and of comparable length, the terminal palpomere longer than the others with a club-shaped apex ( Fig. 29 View FIGURES 28–32 ); labial palp short and divided into three parts, the posterior palpomere as long as the anterior and median palpomeres combined, featuring a rounded and dilated apex ( Fig. 29 View FIGURES 28–32 ); ocelli absent, and the compound eyes slightly reduced, dorsal region near the antennal cavities depigmented ( Figs. 28 and 29 View FIGURES 28–32 ). Thorax: pronotum pubescent, with its anterior and posterior margins arched and covered by elongated bristles, the median region intersected by an anteroposterior groove, lateral lobes subtriangular, facing downward and forward, being slightly shifted towards the anterior region of the pronotum in comparison to P. (T.) kereita n. sp. ( Figs. 29 and 32 View FIGURES 28–32 ); metanotal gland poorly developed, confined to the anterior margin of the metanotum, adorned with numerous setae, concealed by the tegmina, and subdivided into left and right lobes, which are separated by a whitish line that extends anterodorsally until reaching the supra-anal plate. Abdomen: tergites pubescent; supra-anal plate subtriangular and pubescent, with a rounded posterior margin covered by long bristles, slightly shorter than the paraprocts ( Fig. 31 View FIGURES 28–32 ); subgenital plate pubescent, with lateral and posterior margins also covered by long bristles, subtriangular-shaped, apex of the posterior margin straight; cerci pubescent and covered by long setae throughout their length, the inner portion of the base covered by globose setae ( Fig. 31 View FIGURES 28–32 ). Tegmina: slightly more sclerotized than those of P. (T.) kereita n. sp., moderately translucent, pubescent, reduced, and lanceolate, partially covering tergites I and II, as well as the anterior portion of tergite III; inner margins not touching; dorsal field intersected by one or two poorly-marked longitudinal veins, while the lateral field is bordered by an also poorly-marked longitudinal vein curving towards the center of the body, posterior portion of the lateral field whitish and thickened ( Figs. 30 and 32 View FIGURES 28–32 ). Legs: pubescent; leg I (31.80 mm) longer than leg II (28.369 mm), femur bearing small forward-pointing denticles on both ventral margins, tibia devoid of tympanum; tibia of legs I and II bearing two ventral apical spurs; tarsomere I of legs I (5.90 mm) and II (4.45 mm) serrated ventrally and longer than tarsomeres II and III combined; right and left legs III missing, but probably present a distribution of spurs and spines similar to that of P. (T.) kereita n. sp. described herein.
Ecological remarks— Mount Suswa, situated within the Rift Valley in Kenya, ranks among the largest and most intricate Quaternary volcanoes in the region ( Fig. 33 View FIGURES 33–36 ). This imposing geological formation ascends from the southward-sloping Rift Valley floor, assuming the form of an asymmetric, shield-shaped cone with an expansive coverage spanning approximately 270 square kilometers ( Johnson 1969). Comprising mainly sodalite-bearing, phonolitic lava flows along with lesser amounts of pyroclastic materials, the volcanic cone was established atop a preexisting volcanic plateau consisting of quartz-bearing trachyte flood lavas ( Johnson 1969). Mount Suswa is home to one of the most significant lava tube systems in Kenya. Within an approximately 3 square kilometer area, extensive phonolitic basalt flows have formed a complex network of lava tube caves, making it the largest concentration of such caves in East Africa ( Simons 1998a). These caves, numbering around 40 in total, are part of a highly intricate system with multiple levels of passages, spanning an estimated combined length of over 11 kilometers. Most of the cave entrances are the outcome of substantial roof collapses. Notably, there are three distinct levels of passages within this system. The uppermost passages are situated just beneath the lava’s surface crust, whereas the lowest passages can be found at depths ranging from 20 to 35 meters below ground level. It is common to observe passages from different levels overlapping one another. In examining cross-sections of completely filled-in tubes visible in the cave walls, it becomes evident that the network was once even more extensive. The size of these passages varies significantly. In the largest cave systems, passages typically measure between 6 to 10 meters in diameter, although they can expand to 20 to 30 meters where breakdowns have occurred. Furthermore, individual passage segments, extending both from the main chambers and between collapse sites, can span anywhere from 10 meters to over half a kilometer in length ( Simons 1998a).
The Baboons Cave ( Fig. 34 View FIGURES 33–36 ) represents just a small portion of the intricate network of lava tube caves found in the region. These caves underwent an interesting history of human use, some of which have had significant impacts on them. Several decades ago (from 1966 to 1984), these caves were exploited for bat guano, which was used as a fertilizer ( Simons 1998b). The primary producers of this guano were the African Giant Free-Tailed Bats, Otomops harrisoni ( Fig. 35 View FIGURES 33–36 ) ( Ralph et al. 2015). Remarkably, these bats still inhabit the cave system and continue to generate substantial quantities of guano, which accumulates on the cave floor.
Specimens of P. (T.) papiophagus n. sp. were seldom encountered within the cave system. It is worth noting that Simons (1998b) had previously made mention of the presence of “pale crickets with long antennae,” which likely corresponds to the species described herein, or, eventually, to P. (Araneacris) troglophila , which is also present in this cave. Notably, the behavior of P. (T.) papiophagus n. sp. differs markedly from that observed in P. (T.) kereita n. sp. While the latter species displayed relative calmness, P. (T.) papiophagus n. sp. is exceptionally elusive. Individuals were consistently seen in isolation from one another and exhibited a rapid retreat response, even when humans were at a considerable distance (over 3 meters). Given the abundance of rock blocks scattered across the cave floor, capturing these crickets proved to be an extremely challenging task, as they swiftly sought refuge among the rocky terrain. Consequently, despite efforts, only a single adult male specimen was collected. While this hypothesis remains somewhat speculative, this elusive behavior may have been intensified by historical guano removal activities within the cave. These activities likely caused significant disruption to the cricket population. It is important to note that, in addition to guano, these crickets have been observed consuming unconventional organic resources, such as baboon carcasses ( Fig. 36 View FIGURES 33–36 ). Specimens of P. (T.) papiophagus n. sp. were observed feeding on such cadavers, although they swiftly retreated before photographic documentation could be obtained.
The cave system is presently utilized for tourism; however, it lacks infrastructure to support visitors adequately. Moreover, owing to the substantial bat colony within, there exists a potential risk of an epidemic outbreak. Several distinct viruses have previously been documented in Kenyan caves, including those within the Mount Suswa area ( Conrardy et al. 2014; Kamau et al. 2022). Therefore, exercising caution is strongly advised when visiting these caves, and monitoring programs should be instituted to prevent potential epidemic or pandemic outbreaks.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Grylloidea |
|
Family |
|
|
Tribe |
Phalangopsini |
|
SubTribe |
Phaeophilacridina |
|
Genus |
|
|
SubGenus |
Phaeophilacris |