
Ia io peninsulata, Soisook, Pipat, Sribuarod, Kriangsak, Karapan, Sunate, Safoowong, Mongkol, Billasoy, Sawwalak, Thong, Vu Dinh, Chang, Yang, Gong, Lixin, Lin, Aiqing, Sztencel-Jabłonka, Anna, Bogdanowicz, Wiesław & Bates, Paul J. J., 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4344.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:9D1C0264-9C5F-441B-A268-27ADD4E6208B |
|
DOI |
https://doi.org/10.5281/zenodo.6048896 |
|
persistent identifier |
https://treatment.plazi.org/id/03E98782-FFAE-AF20-D2EE-080AC3327182 |
|
treatment provided by |
Plazi |
|
scientific name |
Ia io peninsulata |
| status |
subsp. nov. |
Ia io peninsulata View in CoL subsp. nov.
( Figs. 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 , Tables 1–2)
Holotype: PSUZC-MM.2012.158, adult male, dry skin, skull and baculum extracted, collected by Kriangsak Sribuarod, on 7 July 2012.
Type locality. Tham Lot, Khao Sok National Park, Surat Thani Province, S. Thailand (8°58′N, 98°35′E).
Paratypes. PSUZC-MM.2012.159, adult female, dry skin, skull extracted, from the same site as the holotype, collected by Kriangsak Sribuarod; uncatalogued specimen, field number PS170204.1, adult female, body in alcohol, skull extracted, collected from Panturat Cave, Surat Thani Province, S. Thailand (8°54′N, 98°31′E) by P. Soisook; PSUZC-MM.2012.157, adult male and PSUZC-MM.2012.207, adult female, dry skin, skull extracted, collected from Sra Manora Forest Park, Phang Nga Province, S. Thailand (8°30′N, 98°32′E) by Kriangsak Sribuarod.
Referred specimens. HZM 7.40229 (012/12), cranium and mandible, with almost complete dentition (missing left upper I2, I3, C1, P2; right upper I3, P2; both left and right lower I1, I2, I3); HZM 8.40230 (013/12), cranium only, with almost complete dentition (missing upper left I2, I3; upper right I2, I3, C1, P2); HZM 6.40228 (016/12), mandible only, with almost complete dentition (missing lower left I1, I2, I3, C1, P2) – all three specimens collected in 2012 by Helmut Steiner from the floor of Panturat Cave, Surat Thani Province, S. Thailand (8°54′N, 98°31′E).
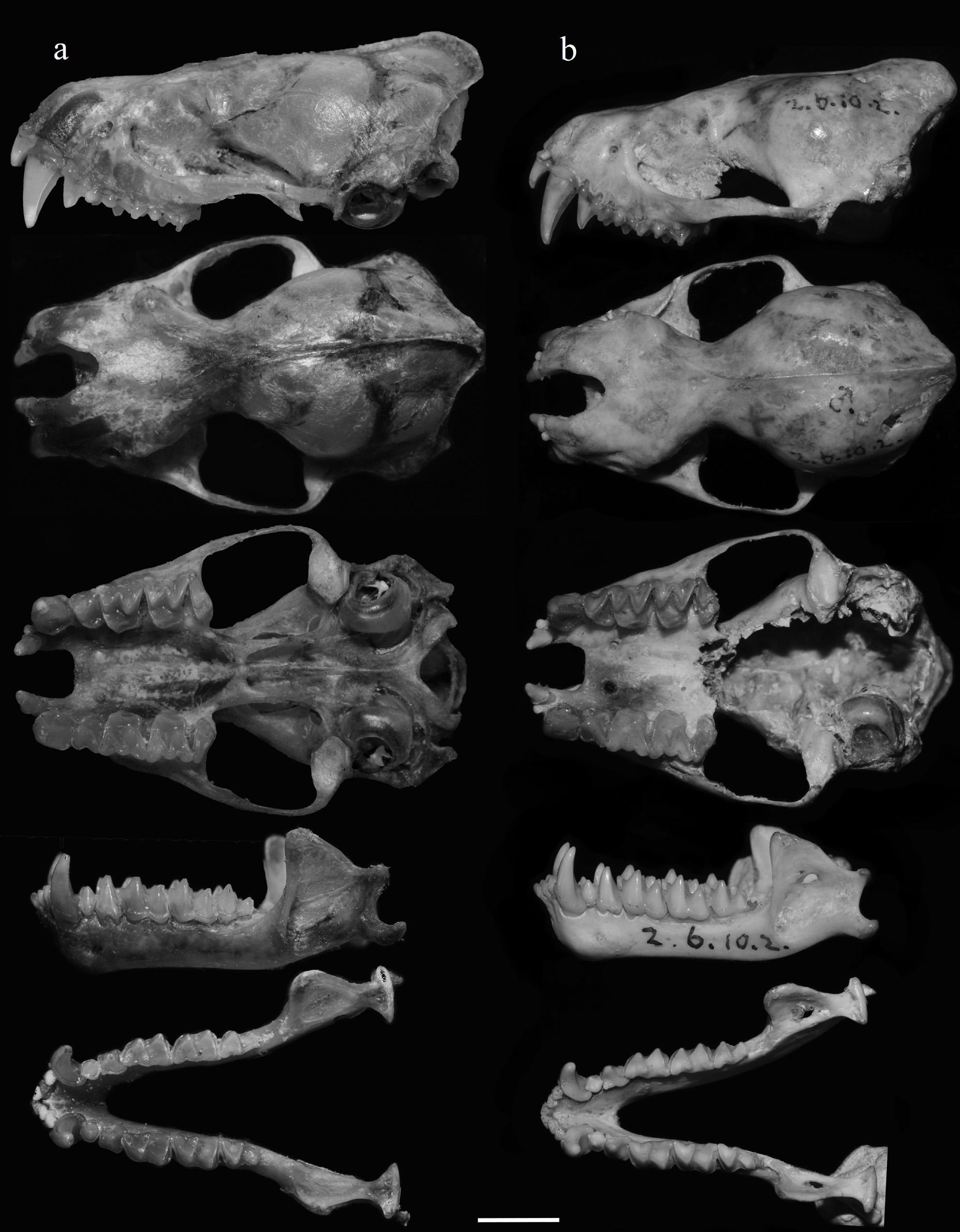
Diagnosis. This is a very large vespertilionid, with a FA of 78.4–83.1 mm ( Table 1). The dorsal pelage is dark brown and the ventral pelage is greyish-brown with an apparent whitish-brown ring on the chest between the shoulders ( Fig. 2 View FIGURE 2 ). The skull is massive ( Fig. 3a View FIGURE 3 ) with a GTL and CCL of 29.30–31.13 and 27.12–28.70 mm, respectively ( Table 1). The inner upper incisor (I1) is unicuspid. The upper canine (C1) is large, more than twice that of the second upper premolar (P4) ( Fig. 3a View FIGURE 3 ). The first lower premolar (P2) is rounded; the second lower premolar (P4) is somewhat square ( Fig. 3a View FIGURE 3 ). The anterior basal cusps of both P2 and P4 are poorly developed. The talonids of the first (M1) and second lower molars (M2) are about equal to that of their respective trigonids ( Fig. 3a View FIGURE 3 ).
Characters Ia io peninsulata subsp. nov. Ia io io P -value
n mean ± SD min–max n mean ± SD min–max
Etymology. The subspecific name refers to the Thai Peninsula, from where specimens of the new subspecies were collected.
Description. This is a very large bat, and probably the largest known vespertilionid, with a FA of 78.4–83.1 mm and HB of 101.2–106.4 mm ( Table 1). The ears are relatively large ( Fig. 2 View FIGURE 2 ), with an E of 25.2–26.7 mm, and a rounded tip. The anterior border of the pinna is rounded, and the anterior rim is thickened; the posterior border is somewhat wrinkled. The tragus is typical and short, with TRG of 10.0– 11.3 mm, and slightly projected anteriorly. The dorsal pelage is dark brown and the ventral pelage is greyish-brown with an apparent whitish-brown ring across the chest ( Fig. 2 View FIGURE 2 ). The hairs on the lower part of belly, the flanks and on those parts of the wings that are closest to the body are somewhat orange brown. The wing and tail membranes are dark brown. The wing membrane attaches to the middle of the outer metatarsus of each foot. The tail is long, with TL of 62.8–74.5 mm. The foot is almost naked, except at the base of the claw where there are some very short hairs. The foot length is about half that of the tibia; HF and TIB are 15.0–18.0 mm and 29.9–36.4 mm, respectively.
The skull is robust, with a GTL and CCL of 29.30–31.13 mm and 27.12–28.70 mm, respectively ( Table 1). The inner upper incisor (I1) is unicuspid and relatively large, about one thirds that of the upper canine (C1) in height ( Fig. 3 View FIGURE 3 ). The outer incisor (I2), however, is very small with the height only about one third that of the I1. The C1 is large, more than twice that of the second upper premolar (P4) ( Fig. 3a View FIGURE 3 ). The first lower premolar (P2) is rounded; the second lower premolar (P4) is somewhat square in outline ( Fig. 3a View FIGURE 3 ). The anterior basal cusps of both P2 and P4 are poorly developed. The talonids of the first (M1) and second lower molars (M2) are about equal to that of their respective trigonids ( Fig. 3a View FIGURE 3 ).
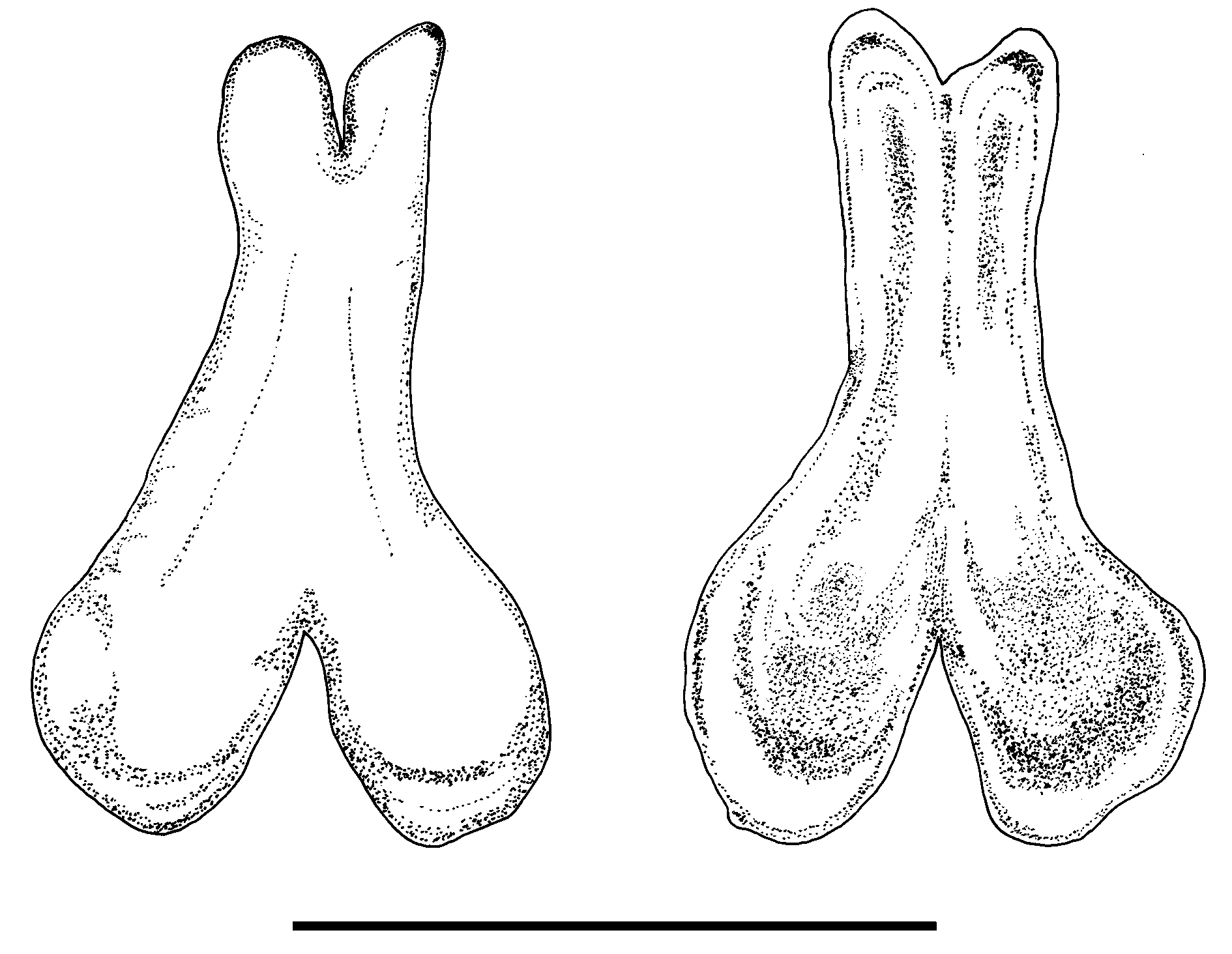
The baculum is small, 1.2 mm in length, with a somewhat unbalanced X-shape ( Fig. 4 View FIGURE 4 ) and with the tip much narrower than the basal end. There is a deep median notch at the tip and a more pronounced one at the base.
Comparisons. The new subspecies, peninsulata, has a larger body size than the nominate form, io (mean FA 80.4 vs 75.2 mm; Table 1). The hair of peninsulata is uniformly dark brown on both the dorsal and ventral sides with a distinct whitish collar over the chest between the shoulders, not observed in io. The skull of peninsulata is more heavily built ( Fig. 3 View FIGURE 3 ), and larger than that of io ( Table 1). For example, the GTL and C1–M3 of peninsulata are significantly larger and do not overlap with io ( Table 1; Fig. 5 View FIGURE 5 ). The upper canine (C1) of peninsulata is massive and subequal to that of the first upper molar (M1) in crown area, whereas it is only about two-thirds in io. The height of the C1 of peninsulata is more than twice that of the P4 but it is only about twice that of the P 4 in io ( Fig. 3 View FIGURE 3 ). In peninsulata, the crown area of the first lower premolar (P2) is larger, more than half that of the second (P4), but in io the crown area of P2 is less than half that of the P4 ( Fig. 3 View FIGURE 3 ). The second lower premolar (P4) is somewhat square rather than rectangular in io ( Fig. 3 View FIGURE 3 ). According to the PCA based on the correlation matrix of 11 craniodental measurements of 19 specimens, the first two components (PC1 and PC2) explain 92.85% of total variation ( Table 2), and it clearly separates specimens of io and peninsulata into two different groups ( Fig. 6 View FIGURE 6 ). The baculum of the new subspecies is smaller and looks substantially different from the specimen of io from India as described and illustrated in Topál (1970), which is 1.8 mm in length and somewhat triangular in shape without median notch at the tip (see figure 2, Topál 1970). The baculum of a specimen from Chiang Mai, northern Thailand (CDWRS-B- 087) is without median notch at the tip and very similar to the specimen described in Topál (1970).
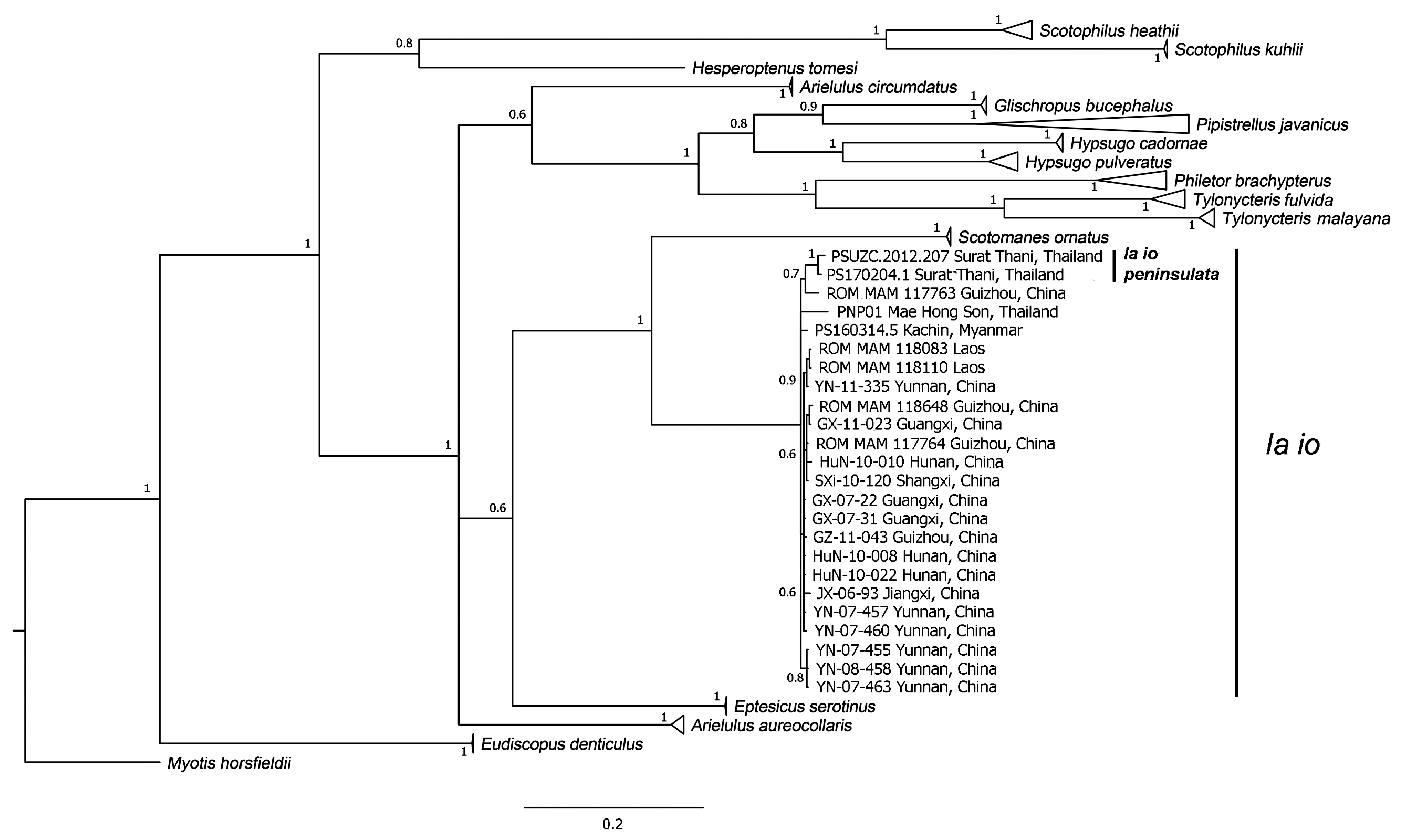
Genetics. Among other vespertilionids, I. io is a sister species of S. ornatus ( Fig. 7 View FIGURE 7 ). The genetic distance based on COI gene between the two species is 18.85%. The new subspecies peninsulata clusters together in a monophyletic clade with specimens of io from China, Laos, Myanmar and northern Thailand ( Fig. 7 View FIGURE 7 ). The pairwise genetic distance between the two subspecies is relatively low, 1.89%, whereas the overall mean distance between I. io specimens analysed in this study is 2.6% based on COI. The topology of the phylogenetic tree based on the preliminary analysis of CytB gene (with only one specimen of the new subspecies available) is very similar to that of the COI, but with a distance between the two subspecies of 1.65%. Based on substitution rate, the divergence time can be estimated at approximately 1.2 million years ago ( Thong et al. 2012) or during one of the Pleistocene glacial periods. Further genetic analysis with additional samples and nuclear markers for a more accurate divergence time is currently being carried out (P. Soisook, unpublished data).
Echolocation. Ia i. peninsulata subsp. nov. emits a multi-harmonic frequency modulated (FM) call. Based on the calls recorded from two individuals (one male and one female), the call comprises four harmonics and the maximum energy is found in the first (bottom) harmonic with an fmaxe of 23.6–27.4 kHz. The start frequency (sf) and terminal frequency (tf) are of 76.5–92.3 and 11.8–14.2 kHz, respectively. In terms of frequency, it is comparable with the recording of a hand-released individual of I. io from Kachin, Upper Myanmar, that emitted FM calls with a sf and tf of 80.9–82.9 kHz, and 14.9–16.3 kHz, respectively; the famxe was 26.0–33.9 kHz, and was also found in the bottom harmonic. However, the call of the individual from Myanmar comprises three harmonics (P. Soisook, unpublished data).
Ecology and distribution. At Panturat Cave, during a day visit by PS, four individuals of I. i. peninsulata were found. Three were hanging together on the cave ceiling at about 4 m above the cave floor, the fourth was hanging singly on the cave wall about 3 m above the cave floor. Ia io shared the roost with over 40 individuals of Megaderma lyra in the inner parts of the cave. The first chamber of the cave was occupied by colonies of approximately 700 individuals of Hipposideros pendleburyi and approximately 200 of Taphozous melanopogon . There were feathers of several bird species found scattered on the cave floor, with at least one identified as the Asian paradise flycatcher ( Terpsiphone paradisi ). It is probable that either M. lyra and/or I. i. peninsulata hunted them since both bat species are known to feed on birds (i.e. Bates & Harrison 1997). However, a preliminary faecal analysis of an individual of I. i. peninsulata from this cave indicated that beetles (Coleoptera) were the preferred prey (over 90% by volume) with no fragment of a vertebrate found (A. Samoh, pers. comm.). At Tham Lot Cave (type locality), a tightly packed group of about 10 individuals of I. i. peninsulata was found roosting together on the cave ceiling about 4 m above the cave floor.
The subspecies peninsulata is currently known from three localities in peninsular Thailand ( Fig. 1 View FIGURE 1 ). This extends the range of I. io southward to the Sundaic Subregion. However, further survey in limestone karst areas may reveal that it is more common and widespread in the Thai-Malay Peninsula.





| HZM |
Museum of Natural History (Hrvatski Zooloski Muzej) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |