
Rafflesia verrucosa Balete, Pelser, Nickrent & Barcelona, 2010
|
publication ID |
https://doi.org/10.11646/phytotaxa.10.1.8 |
|
DOI |
https://doi.org/10.5281/zenodo.4899819 |
|
persistent identifier |
https://treatment.plazi.org/id/03E9357A-E33D-FF9D-A69D-FF7DFB3FFB54 |
|
treatment provided by |
Felipe |
|
scientific name |
Rafflesia verrucosa Balete, Pelser, Nickrent & Barcelona |
| status |
sp. nov. |
Rafflesia verrucosa Balete, Pelser, Nickrent & Barcelona View in CoL , sp. nov. ( Figs. 2A–F View FIGURE 2 , 3A–F View FIGURE 3 )
Rafflesiae baleteae floribus bisexualis, et R. leonardae antheris numerosis, 20-21, etiam R. aurantiae formis ramentorum similis. Autem verrucis prominentibus in perigonio et diaphragmate orae aperturae attingentes, disco cupulato processibus laminaribus sinuosis consociatis confertim impletis, corona pubiscentia densa ad trientum basilarem disci externi extensa, annulo latissimo toroido relative laevi, antheris minimus numerosioris ab omnibus aliis rafflesiis parvis praesertim differt.
Type:— PHILIPPINES: Mindanao Island : Davao Oriental Province: Manay (also known as Man-ay ) Municipality: Mt. Kampalili, 7.29112° N, 126.31520° E, 1470 m, 3 March 2010, Balete 17 ( holotype PNH, isotypes CAHUP, F, L, SING, US) GoogleMaps .
Endophytic holoparasite. Mature buds to 7.5 cm in diameter. Cupule 2.0 cm high to 6.0 cm wide. Bracts (or bud scales) numerous in three to four imbricate whorls, outermost smallest, ca. 1.6 cm long, 1.5 cm wide, innermost largest to ca. 5.1 cm long, 6.5 cm wide. Flowers 14.5–16 cm in diameter when fully expanded, 11.5–13 cm high ( Fig. 2A View FIGURE 2 ), with a very mild putrescent smell when fresh. Perigone tube ca. 3.5–5.5 cm long, inner surface with minute pustules (broccoli-like), with a midlayer of sclerenchymatous tissue that extends to the perigone lobes, this persisting beyond senescence and into fruit maturity, often forming a cup-like structure on top of the maturing fruit ( Fig. 2B View FIGURE 2 ). Perigone lobes 5, orbicular to broadly orbicular, 4.2–4.5 cm long, 6.5–8.0 cm wide, margins irregularly sinuate, reddish orange or cinnamon, becoming dark brown with age; adaxial surface verrucose, warts prominently raised, solitary, irregular in shape, size, and density, usually roundish, less often rod-like to narrowly elongated, white-tipped in newly opened flowers, with age becoming concolorous with perigone adaxial surface, abscising upon flower senescence leaving a persistent wood-like tissue extending from the perigone tube ( Fig. 2B View FIGURE 2 ); abaxial surface covered with hardened tissue that is irregularly cracked and partially flakes off (sometimes already early in the development of the flower) revealing a pitted inner layer ( Fig. 2D View FIGURE 2 ). Diaphragm 7–9 cm in diameter, 2–3.5 cm wide from aperture rim to base of perigone lobes, 9–11 mm thick at base, becoming thinner towards the aperture; concolorous with or slightly darker than the adaxial perigone surface; densely covered with prominently raised, pleated, plate-like warts that are white-tipped in newly opened flowers, these variable in size, larger ones 3–8 mm tall, intermixed with smaller ones, ca. 1–2 mm tall ( Fig. 2C View FIGURE 2 ); windows absent; aperture 3.5–4 cm in diameter. Disk 5–5.5 cm in diameter, ca. 1–1.2 cm thick midway between the margin and the point where the disk joins the column, cup-shaped with a prominently raised margin, concolorous with diaphragm; processes forming an interconnected system of tightly packed, laminar plates with erose margins, ca. 3–11 mm tall ( Fig. 2C, E–F View FIGURE 2 ); column very short, up to ca. 1 cm from the floor of the floral tube to the upper surface of disk, ca. 2.3 cm wide, deeply grooved, number of sulci corresponds to the number of anthers, septa between sulci ca. 1 cm tall, with acicular hairs (‘bristles’ sensu Nais 2001) ca. 1.5 mm long; annulus ca. 7–10 mm wide, ca. 5–6 cm in diameter, doughnut-shaped (toroid), concolorous with the rest of the perigone tube, surface shallowly pitted and rugose ( Fig. 2F View FIGURE 2 ). Ramenta to 7 mm long, covered with clavate pustules, polymorphic, filiform to variously branched or cleaved apically, those at the floor of the perigone tube longer and denser becoming shorter and more widely spaced on the diaphragm, nearly absent near the aperture rim, white-tipped in newly opened flowers, becoming concolorous with perigone abaxial surface with age. Flowers bisexual ( Fig. 3A View FIGURE 3 ); anthers 20 (or 21), globular, ca. 2 mm in diameter, prominently protruding from very shallow anther sulci that are 2–3 mm long, 3–4 mm wide ( Fig. 3B View FIGURE 3 ), basal third of disk formed into a pubescent corona with bristle-like, acicular hairs to 0.5 mm ( Fig. 2E View FIGURE 2 , 3B View FIGURE 3 ); ovary ca. 1.5 cm tall, 4.5 cm wide, lenticular or reniform, becoming broadly and irregularly so as the ovary matures ( Fig. 3C View FIGURE 3 ); developing fruits to 7 cm in diameter and 4.7 cm high, sclerenchymatous perigone tissue persistent.
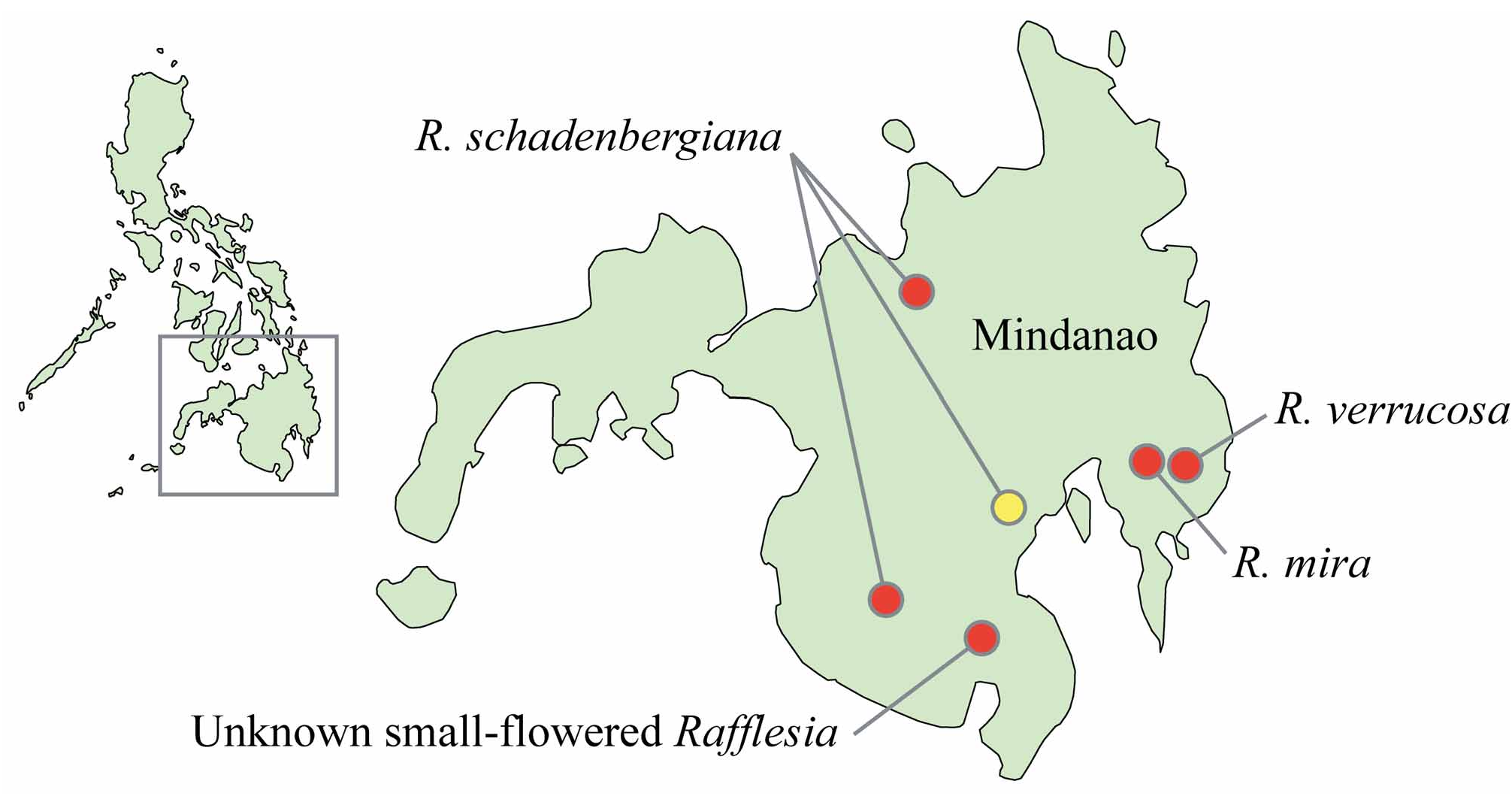
Distribution and habitat:— Rafflesia verrucosa is only known from the southeastern slope of Mt. Kampalili, Davao Oriental Province, Mindanao Island ( Fig. 1 View FIGURE 1 ). It is found between 1350 and 1550 m. Despite extensive search efforts in the area, it was not encountered at lower ( 900–1350 m) or higher ( 1550–1700 m) elevation, although the presence of Tetrastigma vines, which is the host for this species, was confirmed throughout the elevational range explored.
The habitat of R. verrucosa is montane forest in relatively rocky and moderately sloping terrain. Leaf litter is ca. 2–5 cm thick. The canopy is ca. 15 m tall and emergents reach 20 m. The larger trees in this forest commonly reach a DBH up to ca. 90 cm. Dominant trees are oaks ( Lithocarpus Blume 1826: 526 ), laurels ( Litsea Lamarck 1792: 574 ), and myrtles ( Syzygium Gaertner 1788: 166 ). Mosses are common on tree trunks and branches, and often also at the bases of trunks. Other epiphytes include ferns and lycophytes, Medinilla Gaudichaud-Beaupré (1830: 484) , and orange-flowered Rhododendron Linnaeus (1753a: 392) . Understory vegetation is abundant and composed of tree ferns, other ferns and lycophytes, ground orchids, gingers and grasses. Canopy vines such as Freycinetia Gaudichaud-Beaupré (1824: 509) , climbing bamboos, rattans, Smilax Linnaeus (1753b: 1028) , Tetrastigma , and other lianas are likewise common. Several R. verrucosa plants were found on Tetrastigma vine roots growing exposed on rocky ground which resulted in deformed buds and flowers.
Etymology:— The specific epithet of this new taxon is derived from the Latin verruca (wart), which calls attention to the unique, prominently raised warts on the perigone lobes and diaphragm.
Ecology:— Rafflesia verrucosa is restricted to plants of a single, presently unidentified species of Tetrastigma ( Fig. 3D View FIGURE 3 , specimen not collected). Flowers and buds were only observed on the roots of their host plants and are absent on prostrate and aerial stems. Tetrastigma roots bearing Rafflesia flowers or scars range from 6 to at least 25.5 mm in diameter and buds were found on roots buried up to 7 cm below ground level.
A total of 15 clusters of flowers and buds were discovered in an area of ca. 20 m × 1 km. At one site, at least 30 Rafflesia buds in different stages of development as well as senescent flowers were observed ( Fig. 3E View FIGURE 3 ). This is equivalent to ca. 7–8 flower clusters per hectare. At the time of discovery of R. verrucosa (February–March, 2010), a notably large number of senescent flowers and developing fruits were observed. This suggests that flowering must have peaked around October to December similar to what is reported for R. mira , a species found in the adjacent Mt. Candalaga, Compostela Valley ( Madulid et al. 2006). Since some mature buds, newly opened flowers, and early senescent flowers were also observed during our visit in February–March, flowering appears to continue intermittently throughout the year.
Several buds as well as a maturing ovary of a senescent flower showed evidence of damage by animals, mainly as bite marks of unidentified small-toothed mammals ( Fig. 3F View FIGURE 3 ). Systematic trapping in the area where R. verrucosa occurs yielded at least ten species of native non-volant small mammals, including a gymnure ( Podogymnura ), a shrew ( Crocidura beatus ), a tree shrew ( Urogale everetti ), two shrew-mice ( Crunomys ), a moss-mouse ( Tarsomys ), an arboreal tree-mouse ( Apomys ), a forest mouse ( Apomys ), the large Mindanao forest rat ( Bullimus bagobus ) and the common Philippine forest rat ( Rattus everetti ). Also reported in the area by local hunters are two species of squirrels ( Exilisciurus concinnus and Sundasciurus philippinensis ). Likewise, we also documented the presence of larger mammals, including palm civet ( Paradoxurus hermaphroditus ), Philippine warty pig ( Sus philippinensis ) and Philippine brown deer ( Cervus mariannus ). At ca. 1500 m, wild pig diggings on the ground around three populations of R. verrucosa are extensive but no sign of feeding on the buds and flowers was observed, although several were trampled upon. The role of these mammals in Rafflesia dispersal in the Philippines remains unstudied. But in Borneo, for instance, both squirrels ( Callosciurus ) and tree shrew ( Tupaia ) are known to feed on Rafflesia fruits ( Emmons et al. 1991).
Conservation:— Rafflesia verrucosa plants grow in the tropical montane forest formation ( Fernando et al. 2008) which is increasingly undergoing disturbance and fragmentation from clearing for abaca ( Musa textilis Née 1801: 123 ) plantations by native Mandayas. In addition, trees in the area are cut to harvest the young palm heart of the rattan plants that climb these trees. This practice resulted in areas of up to 100 m 2 in which tall trees have been cleared and the understory has been damaged. Despite this disturbance, the forest of the Mt. Kampalili Range is relatively intact and certainly does not suffer from the same degradation and loss that plague the lowland forest in which the majority of the Philippine Rafflesia are found ( Barcelona et al. 2009b). Although R. verrucosa is moderately common in the area, it is not known from any other site on Mt. Kampalili or elsewhere and we therefore recommend that the clearing of the forest for abaca plantations and the harvesting of other forest products must be carefully managed in the wider area to ensure the continued survival of this new species of Rafflesia . We strongly support efforts by local people (e.g. Taocanga Tribal Council Association), environmental organizations (e.g. Philippine Eagle Foundation) and concerned government agencies (e.g. Department of Environment and Natural Resources and the Department of Tourism Region XI) to place certain portions of Mt. Kampalili under protected area and ancestral domain management.
Additional specimens examined ( paratypes):— PHILIPPINES: Mindanao Island: Davao Oriental Prov., Manay (= Man-ay ) Municipality : Mt. Kampalili , 7.29796°N, 126.31216°E, ca. 1378 m, 25 February 2010, GoogleMaps Balete 16 ( SING); 7.29537°N; 126.31606°E, ca. 1550 m, 1 March 2010, Balete 18 ( PUH, US). GoogleMaps


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |