
Balaenoptera bertae n.
|
publication ID |
https://doi.org/ 10.5252/g2013n4a5 |
|
DOI |
https://doi.org/10.5281/zenodo.4818610 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7DD69-FFC5-867B-2466-798FFE2E3F51 |
|
treatment provided by |
Felipe |
|
scientific name |
Balaenoptera bertae n. |
| status |
n. |
Balaenoptera bertae n. View in CoL sp.
( Figs 11-15 View FIG View FIG View FIG View FIG View FIG )
HOLOTYPE. — UCMP 219078 View Materials , a partial skull lacking the premaxillae, maxillae, nasals, jugals, tympanic bullae, and bodies of the petrosal, excavated by R. W. Boessenecker,
E. Johnson, T. Palladino, and M. Berrini from UCMP locality V99849 View Materials from August 12-15, 2005.
TENTATIVELY REFERRED SPECIMENS. — UCMP 131815, a partial juvenile cranium including the vertex, supraorbital processes of the frontals, palatines, and tips of the zygomatic processes collected in 1985 by a UCMP party from UCMP locality V85027 View Materials ; UCMP uncataloged, an isolated squamosal from UCMP locality V99868 View Materials .
ETYMOLOGY. — Honouring Dr Annalisa Berta (San Diego State University), for her many contributions to the study of fossil cetaceans and pinnipeds.
DIAGNOSIS. — Balaenoptera bertae n. sp. is a small balaenopterid (estimated bizygomatic width of 61.4 cm), smaller in size than extant Balaenoptera acutorostrata Lacépède, 1804 , characterized by: broadly triangular occipital shield that laterally overhangs the temporal fossa and exhibits a flattened apex, large occipital condyles, little to no parietal exposed dorsally at the transversely narrow vertex, squamosal with short, anteriorly directed zygomatic process, posteriorly elongate postglenoid process, prominent squamosal crease, frontal with rectangular supraorbital process with transversely oriented anterior margin, frontal that is abruptly depressed below the vertex and obliquely oriented so that this surface and anterior wing of parietal are visible in dorsal aspect, large petrosal fossa (sensu Mead & Fordyce 2009), and a posterior process of the petrotympanic that is relatively short and inflated.
STRATIGRAPHIC OCCURRENCE. — Uppermost part of the San Gregorio section of the Purisima Formation (sensu Powell et al. 2007) that is bracketed below by an ash bed correlated with the 3.3-3.4 Ma Putah Tuff (Powell et al. 2007) and above by another ash bed correlated with the 2.5 ± 0.2 Ma Ishi Tuff. Ŋis indicates an age of 3.35-2.5 Ma, or Early to Late Pliocene (Piacenzian- Gelasian equivalent).
TYPE LOCALITY. — UCMP locality V99849 View Materials , Pliocene Purisima Formation. Detailed locality information available on request from UCMP or the author.
DIFFERENTIAL DIAGNOSIS
Balaenoptera bertae n. sp. is a member of the extant genus Balaenoptera and shares with all extant species of Balaenoptera a squamosal crease, relatively short and anteriorly oriented zygomatic processes, a bulge at the squamosal-parietal suture in the posterior temporal fossa, and a truncated apex of the occipital shield.
Balaenoptera bertae n. sp. differs from:
– Archaebalaenoptera castriarquati Bisconti, 2007 , in having a broader occipital shield with a laterally sinuous margin that overhangs the temporal fossa and an anteriorly truncated apex, a transversely thinner vertex measuring only 10% of bizygomatic width (compared to 24% bizygomatic width in A.castriarquati ), short and blunt anteriorly directed zygomatic process that lack supramastoid crest that extends to the zygomatic apex, and having an anteroposteriorly shorter supraorbital process of the frontal;
– “ Balaenoptera ” cortesi “var.” portisi, in having a squamosal crease, shorter and more anteriorly oriented zygomatic process that lacks a supramastoid crest that extends to the anteroventrally oriented zygomatic apex, transversely wide postglenoid process, an anteroposteriorly short supraorbital process with a transversely oriented anterior margin and laterally prominent antorbital process, a shallow sternomastoid fossa, and a large petrosal fossa;
– “ Balaenoptera ” ryani Hanna & McLellan, 1924, in having a posteriorly broad occipital shield, little to no parietal exposed at the vertex, and an anteriorly truncated occipital shield;
– Balaenoptera acutorostrata , in its smaller size, apex of the occipital shield narrow and only 10% of bizygomatic width (25% bizygomatic width in B.acutorostrata ), lacking a lateral triangular extension of the parietal between the squamosal and nuchal crest, exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, transversely oriented anterior margin of the supraorbital process of the frontal in dorsal aspect (as opposed to being posterolaterally oriented), and a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera bonaerensis Burmeister, 1867 , in its smaller size, apex of the occipital shield narrow and only 10% of bizygomatic width (22% bizygomatic width in B. bonaerensis ), exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, and transversely oriented anterior margin of the supraorbital process of the frontal in dorsal aspect (as opposed to being posterolaterally oriented), and a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera borealis Lesson, 1824 , in its much smaller size, apex of the occipital shield narrow and only 10% of bizygomatic width (26% bizygomatic width in B. borealis ), exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, shorter and blunt zygomatic process of the squamosal, shallower squamosal crease, a more posteriorly projecting postglenoid process, a supraorbital process of the frontal that lacks a dorsal ridge, with a more concave lateral margin and transversely oriented anterior margin, larger occipital condyles, a much larger petrosal fossa, and a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera edeni Anderson, 1879 , in its much smaller size, apex of the occipital shield narrow and only 10% of bizygomatic width (18% bizygomatic width in B. edeni ), larger occipital condyles, shorter zygomatic process of the squamosals, a more laterally prominent preorbital process, exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, supraorbital process of the frontal that lacks a dorsal ridge, and a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera musculus Linneaus, 1758 , in its much smaller size, in having a supraoccipital that is more broadly triangular with horizontal (rather than dorsolaterally) nuchal crests that dorsally overhang the temporal fossa, apex of the occipital shield narrow and only 10% of bizygomatic width (22% bizygomatic width in B. musculus ), exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, shorter zygomatic process of the squamosal that is anteriorly directed (rather than anterolaterally directed), larger petrosal fossa and occipital condyles, posteriorly directed postglenoid process (not posterolaterally directed as in B. musculus ), a supraorbital process of the frontal with transversely oriented anterior margin and posterolaterally oriented posterior margin (as opposed to posterolaterally and posteromedially, as in B. musculus ) and a blunt distal apex of the inflated compound posterior process of the petrotympanic; – Balaenoptera omurai Wada, Oishi & Yamada, 2003 , in its smaller size, in having a shorter and blunt zygomatic process of the squamosal, apex of the occipital shield narrow and only 10% of bizygomatic width (24% bizygomatic width in B. omurai ), a more transversely oriented anterior margin of the supraorbital process of the frontal (as opposed to posterolaterally as in B. omurai ), exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, in lacking a sheet of the parietal that medially overlaps the supraorbital process of the frontal, and possessing a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera physalus Linneaus, 1758 , in its much smaller size, apex of the occipital shield narrow and only 10% of bizygomatic width (25% bizygomatic width in Balaenoptera physalus ), exhibiting a ventrolaterally sloping (rather than vertical) surface of the parietal and frontal lateral to the supraorbital process of the frontal, zygomatic process of the squamosal that is shorter and blunt, transversely narrower postglenoid process, a transversely oriented anterior margin of the supraorbital process of the frontal (as opposed to posterolaterally as in B. physalus ), larger petrosal fossa, and a blunt distal apex of the inflated compound posterior process of the petrotympanic;
– Balaenoptera siberi Pilleri & Pilleri, 1989 , in possessing an anteriorly truncated and narrow apex of the occipital shield, only 10% of bizygomatic width (18% bizygomatic width in B. siberi ), a squamosal crease, an anteriorly directed zygomatic process with a supramastoid crest that does not extend to the zygomatic apex, a zygomatic process that is dorsoventrally thicker, a paroccipital process that is less posterolater - ally directed and not as prominent posteriorly, well defined crease between the squamosal and paroccipital process, and a transversely oriented anterior margin of the supraorbital process of the frontal;
– Cetotheriophanes capellinii (Brandt, 1873) , in having a broader apex of the occipital shield that measures 10% of bizygomatic width (4% in C. capellinii ) that is anteriorly truncated, a shorter intertemporal region, a more anteriorly thrusted occipital shield, a shorter zygomatic process of the squamosal, and
A
longer postglenoid processes of the squamosal that are visible in dorsal aspect;
– Diunatans luctoretmurgo Bosselaers & Post, 2010 , in having a narrow apex of the occipital shield measuring only 10% of bizygomatic width (24% bizygomatic width in D. luctoretmurgo ), more sinuous lateral margin of the occipital shield, a slightly narrower zygomatic process that is less laterally offset from the braincase, pterygoid that is ventrally flat and not ventrally rounded in lateral aspect, and a posterior process of the petrotympanic that is anteromedially curved;
– Megaptera novaeangliae Brisson, 1762 View in CoL , in its much smaller size, in having a squamosal crease, shorter and more anteriorly oriented zygomatic process that lack a supramastoid crest that extends to the zygomatic apex, transversely narrow and posteriorly projecting postglenoid process of the squamosal, a less transversely and anteroposteriorly broad supraorbital process of the frontal, a transversely oriented anterior margin of the supraorbital process (as opposed to posterolaterally oriented), anteriorly truncated and narrow apex of the occipital shield measuring only 10% of bizygomatic width (14%
bizygomatic width in M. novaeangliae ), large petrosal fossae, and occipital condyles that are not set out on a distinct neck
– “ Megaptera ” hubachi Dathe, 1983, in having a squamosal crease, shorter and more anteriorly oriented zygomatic process that lacks a supramastoid crest that extends to the zygomatic apex, a more anteriorly thrust occipital shield with a slightly transversely narrower apex measuring 10% of bizygomatic width (compared to 14% in “ M. ” hubachi), an occipital shield that laterally overhangs the temporal fossa to a greater degree, a well defined crease between the paraoccipital process of the exoccipital and the squamosal, transversely narrow and posteriorly elongate postglenoid process of the squamosal, a laterally prominent antorbital process of the frontal, transversely oriented anterior margin of the supraorbital process, an inflated posterior process of the petrotympanic, and by lacking paired tuberosities on the supraoccipital and an anterior indentation in the apex of the occipital shield;
– “ Megaptera ” miocaena Kellogg, 1922, in its smaller size, in having a squamosal crease, shorter and more anteriorly oriented zygomatic process that lacks a supramastoid crest that extends to the zygomatic apex, less anteriorly thrust oc- cipital shield that is roughly triangular and with more widely diverging lateral margins, vertex that is more anteriorly constricted and transversely narrower and measuring 10% of bizygomatic width (compared to 21% in “ M. ” miocaena), an anteroposteriorly broader supraorbital process of the frontal lacking a posterolaterally oriented ridge on its dorsal surface and with a laterally prominent antorbital process, a planar and transversely oriented posterior margin of the supraorbital process of the frontal, a well defined crease between the paroccipital process and squamosal, and an occipital shield that is dorsally convex in lateral aspect;
– Parabalaenoptera baulinensis Zeigler, Chan & Barnes, 1997 , in having a squamosal crease, shorter and more anteriorly oriented zygomatic processes that lack a supramastoid crest that extends to the zygomatic apex, a clearly defined crease between the paroccipital process of the exoccipital and the squamosal, transversely broader apex of the occipital shield that measures 10% of bizygomatic width (8.7% in P. baulinensis ) large occipital condyles, transversely oriented anterior and posterolaterally oriented posterior margin of the supraorbital process of the frontal (posterolaterally and posteromedially oriented in Parabalaenoptera Zeigler, Chan, & Barnes, 1997 , respectively), anterolaterally oriented nuchal crests (oriented dorsolaterally in Parabalaenoptera ), and a transversely narrow and posteriorly elongate postglenoid process of the squamosal;
– Plesiobalaenoptera quarantellii Bisconti, 2010 , in having a less elongate and more inflated and anteromedially curved posterior process of the petrotympanic;
– Protororqualus cuvieri (Fischer, 1829) , in having a squamosal crease, shorter and more anteriorly oriented zygomatic process that lack a supramastoid crest that extends to the zygomatic apex, a supraoccipital apex that is anteriorly squared-off at the vertex and transversely wider, measuring 10% of bizygomatic width (compared to 5% in P. cuvieri ), little to no parietal exposure at the vertex, laterally prominent antorbital process, and a posteriorly elongate postglenoid process of the squamosal.
DESCRIPTION
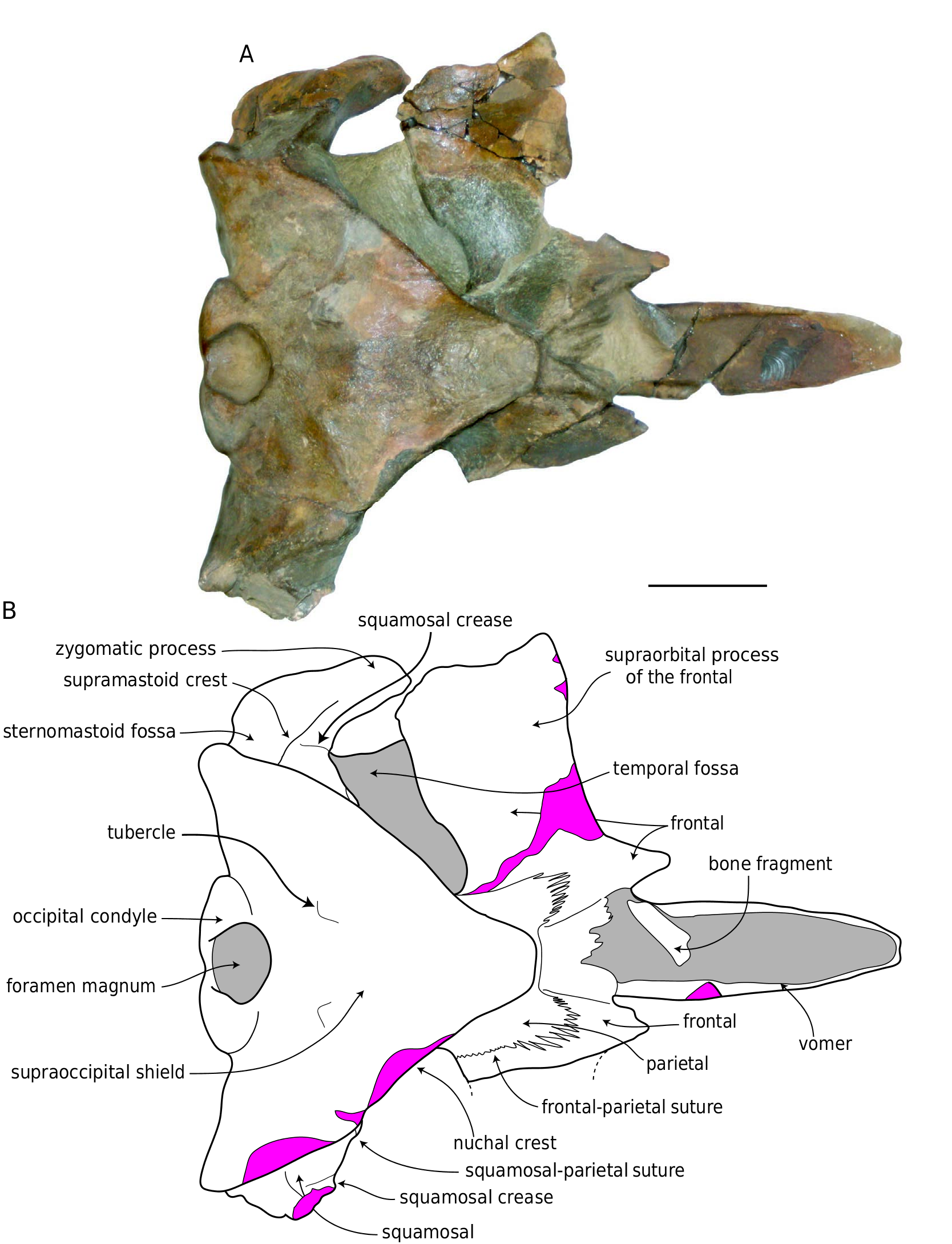
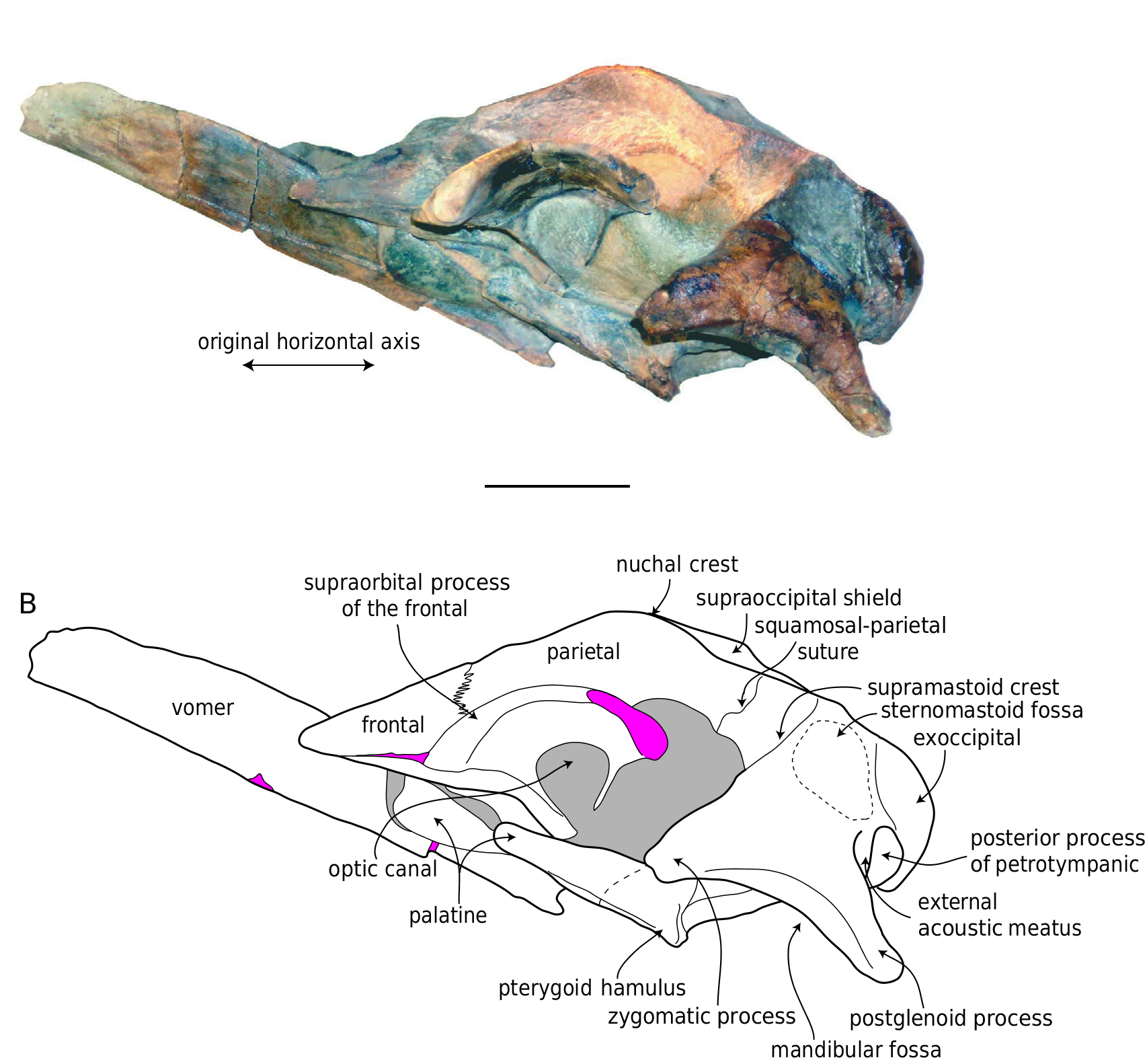
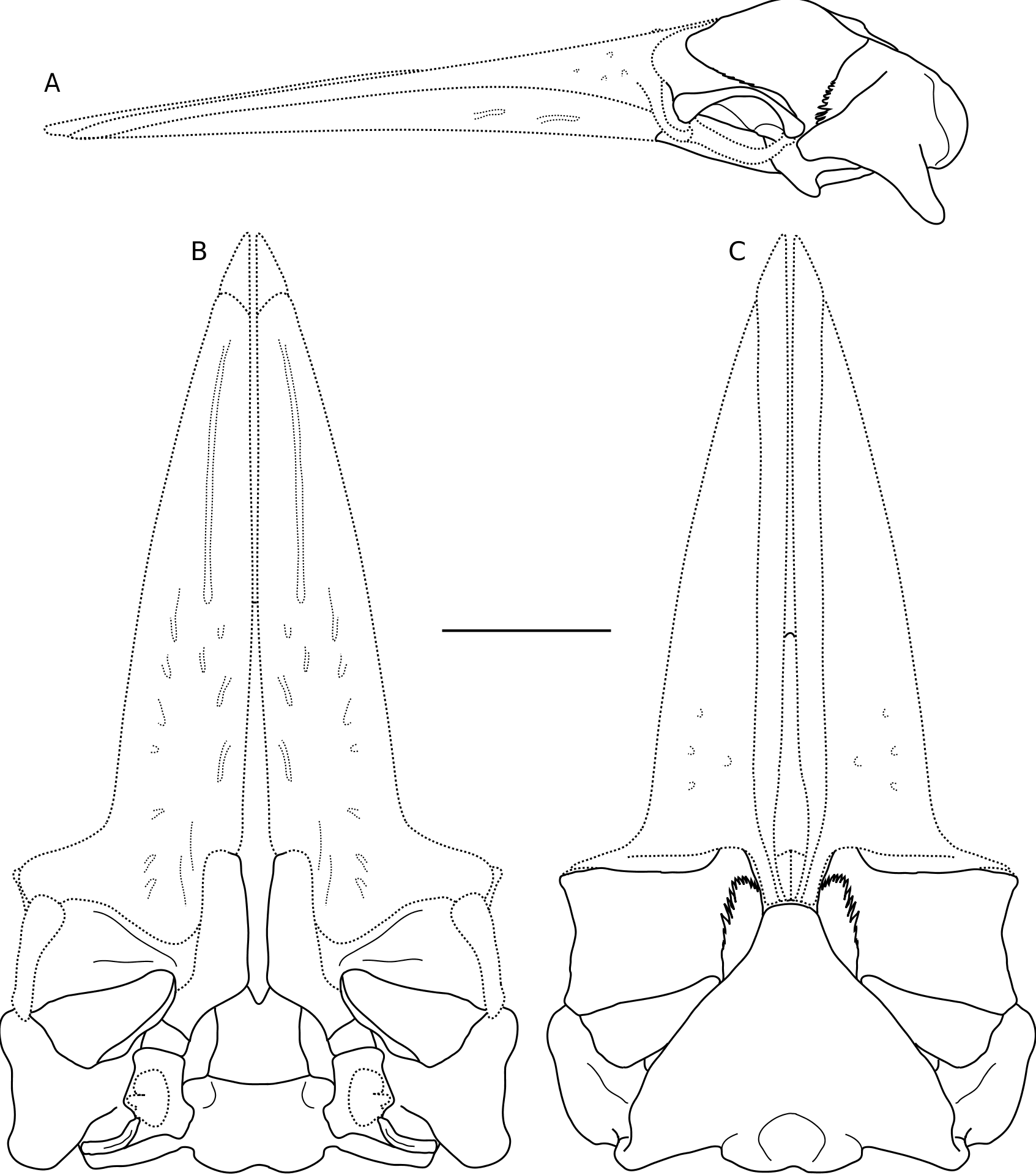
Ŋe holotype is a partial skull including a nearly complete braincase, complete left squamosal and frontal, and vomer; the maxillae, premaxillae, and nasals are missing as well as both right and left tympanic bullae and petrosals (with the exception of the posterior processes; Figs 11-15 View FIG View FIG View FIG View FIG View FIG ). Measurements of the holotype are presented in Table 6 View TABLE . UCMP 219078 View Materials was found upside-down, and many bones of the ventral portion of the skull (as well as the left supraorbital process of the frontal) are fractured and displaced ( Fig. 16 View FIG ). Ŋe vomer is broken posteriorly, and is rotated anterodorsally ( Fig. 13 View FIG ). Ŋe left supraorbital process exhibits a long, anterolaterally oriented fracture, and the portion of the bone lateral to this fracture has rotated so that the supraorbital process is oriented dorsolaterally, and the postorbital process is no longer in contact with the zygomatic process. A reconstruction of the skull of B. bertae n. sp. is depicted in Figure 17 View FIG .
Occipital shield
In dorsal view the occipital shield is broad and triangular with a transversely narrow apex that is rectilinear and transversely oriented in dorsal view ( Figs 11A View FIG , 15A View FIG ). Ŋe anteriormost portion of the shield is longitudinally concave, and posterior to this, it is dorsally strongly convex along the sagittal plane. A pair of faint tubercles occur in the mid- dle of the supraoccipital, anterior to each occipital condyle. Ŋe nuchal crests are formed dorsally by the supraoccipital, are oriented anterolaterally, and overhang the temporal fossa so that the medial wall of the temporal fossa is not visible in dorsal aspect.
Exoccipital
Ŋe posterior margin of the exoccipital is posterolaterally oriented ( Figs 11A View FIG , 12A View FIG ). In dorsal view, the lateral margin of the occipital shield is slightly sinuous. Ŋe occipital condyles are relatively large. Ŋere are no clear dorsal condyloid fossae. Ventral to the foramen magnum, a shallow furrow extends between the occipital condyles.
Squamosal
Ŋe squamosal has a posteriorly elongate postglenoid process that is paddle-shaped (in dorsal and ventral view), dorsoventrally compressed, and posteroventrally directed ( Figs 11B View FIG , 13A View FIG , 15C View FIG ). Ŋe posterior margin of the postglenoid process is semi-circular in dorsal aspect. In lateral aspect, the ventral border of the mandibular fossa is strongly concave ( Fig. 13 View FIG ), and both the zygomatic and postglenoid processes are somewhat ventrally directed, and in dorsal and ventral aspect, the zygomatic process is nearly anteriorly directed ( Figs 11 View FIG , 12 View FIG ). Ŋe zygomatic process is relatively short, blunt, and triangular in crosssection. Ŋe supramastoid crest runs along the dorsal margin of the squamosal from the lateral margin of the occipital shield, but does not extend far onto the zygomatic process. A distinct dorsoventrally oriented and anteriorly facing squamosal crease occurs on the anterior surface of the squamosal, at the base of the zygomatic process. Along the posteromedial wall of the temporal fossa near the parietal-squamosal suture, a faint squamosal cleft occurs and runs parallel to the parietal-squamosal suture. In dorsal aspect, postglenoid process terminates well anterior to the exoccipital. A vertical crease occurs at the squamosal-exoccipital suture at the posterolateral edge of the cranium ( Fig. 11A View FIG ). Ŋe posterolateral surface of the squamosal is relatively planar. Ventrally, the mandibular fossa is transversely broad, and anteroposteriorly concave.
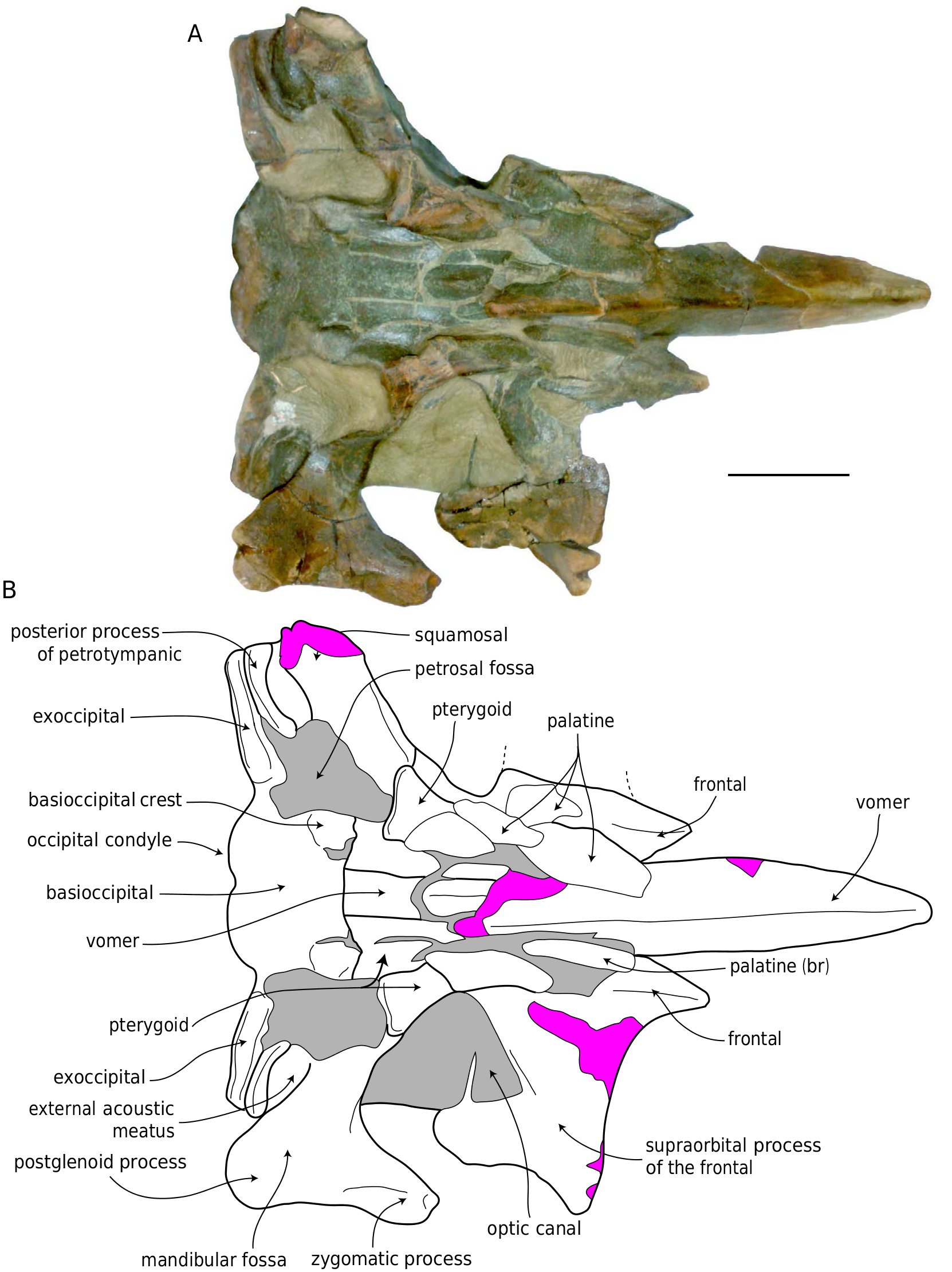
Basicranium
Portions of the basicranium are badly shattered ( Figs 11B View FIG , 12B View FIG ). Ŋe posterior sheet and wings of the vomer and palatines are broken. Ŋe basioccipital is broad and shallowly concave between the relatively small and knoblike basioccipital crests. Ŋe basioccipital crest is elongate and transversely convex. Ŋe petrosal fossa is large, subrectangular, anteriorly bordered by the pterygoid, medially by the medial lamina of the pterygoid, laterally by the squamosal, and posteriorly by the exoccipital ( Figs 11B View FIG , 12B View FIG , 15C View FIG ). Ventrally, the exoccipital forms a transversely oriented ridge that is posterolaterally directed. Posterolaterally, the exoccipital contacts the posterior surface of the posterior process of the petrotympanic. Ŋe posterior process of the petrotympanic curves anteromedially, is distally swollen, and bears a sharp ventral crest ( Fig. 15B, C View FIG ). Ŋe posterior meatal crest anteriorly overlaps the posterior process of the petrotympanic, and anterior to this thin plate, the squamosal exhibits a posterolaterally directed, trough-like external acoustic meatus positioned posterodorsally to the postglenoid process. Ŋe anterior meatal crest continues medially on the squamosal from the postglenoid process and forms the anterior margin of the external acoustic meatus.
Ŋe vomer is nearly complete, and anteriorly protrudes beyond the anterior margin of the frontals ( Figs 11-13 View FIG View FIG View FIG ). Dorsally, the transversely concave mesorostral canal is filled with matrix. Anteriorly,the vomer becomes transversely narrower. Ŋe vomer is V-shaped in cross-section and bears a sharp ventral crest. Posteriorly, the ventral ridge of the vomer becomes lower to merge with the flat wings of the vomer.Ŋe latter becomes relatively wider posteriorly, ventrally flooring the basicranium, contacting the medial lamina of the pterygoid, and underlapping the anterior portion of the basioccipital. Posteriorly, the vomer is broad and flat. Ŋe basisphenoid is not exposed. Ŋe now-fractured palatine appears to have been slightly convex transversely, medially contacting the vomer, and posteriorly contacting the pterygoid. Ŋe palatal surface of the pterygoid is triangular in shape and positioned along the anterior margin of the petrosal fossa, with a blunt, tongue-shaped hamulus. Ŋe pterygoid hamulus does not appear to be broken or to possess fingerlike posteromedial projection as in species of extant Balaenoptera , and this may tentatively considered to be natural. Medially, the blunt pterygoid hamulus overhangs the medial lamina of the pterygoid. Ŋe medial lamina is anteroposteriorly elongate, ventrally flat, and laterally forms the ventrally directed pharyngeal crest that is confluent with the basioccipital crest and forms the medial margin of the petrosal fossa.
Vertex
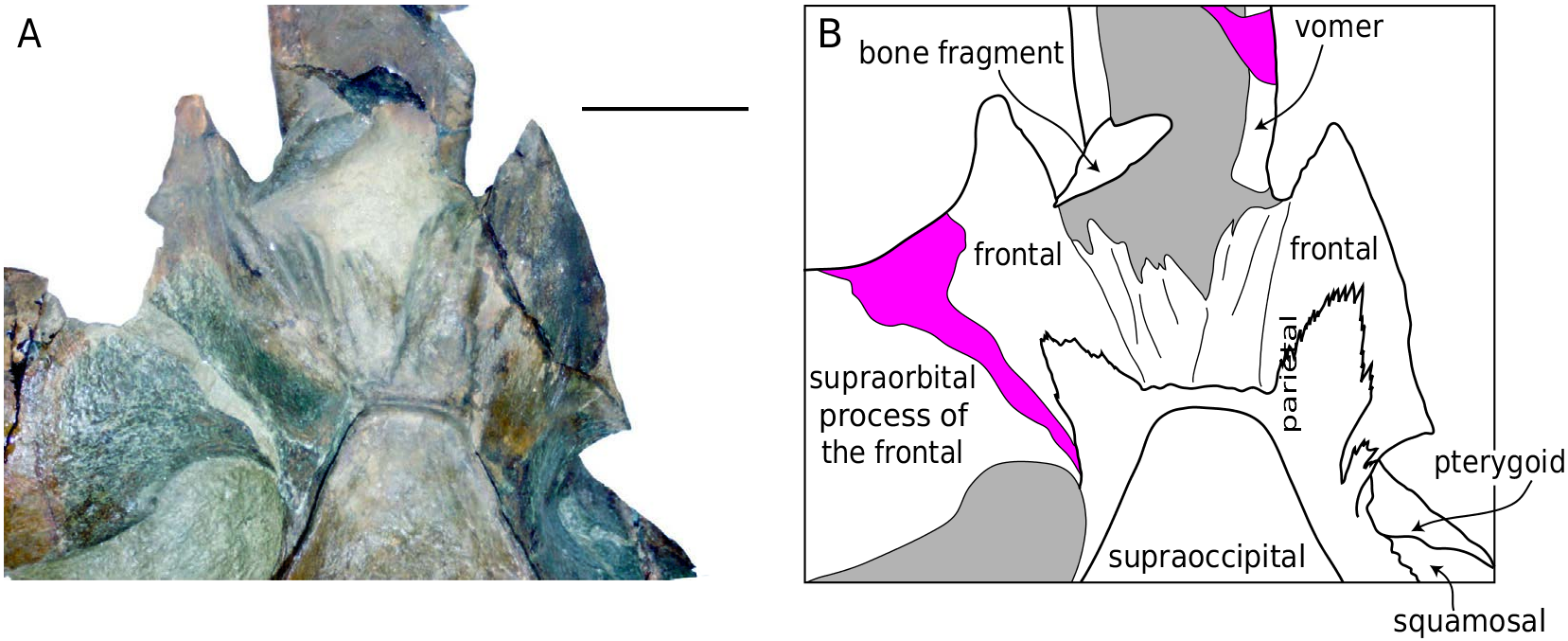
Ŋe vertex of UCMP 219078 is transversely narrow, reflecting the narrow apex of the supraoccipital ( Figs 11A View FIG ; 12A View FIG ; 14 View FIG ). A thin band of the parietal is exposed between the sutures for the rostral elements and the supraoccipital ( Fig. 14 View FIG ). As the rostral elements are missing, it is not possible to ascertain the length of the nasals and morphology of the ascending processes of the maxillae and premaxillae. Ŋe articular surface on the frontals for the rostral elements is corrugated and exhibits a series of anteroposteriorly oriented grooves and ridges; seven ridges are present on the left side; the right side is not as well-preserved. Ŋe region of the vertex as preserved on the frontal appears asymmetrical with the left side having extending further anterolaterally and with a larger angle to the midline; this is due to the slight counterclockwise rotation of the left supraorbital process of the frontal.
Frontal
Ŋe well-preserved left frontal ( Figs 11-13 View FIG View FIG View FIG ) has a sharp, triangular prong anteriorly. Lateral to this prong, the anterior margin of the supraorbital process is straight and transversely oriented in dorsal view ( Fig. 17 View FIG ). Ŋe base of the supraorbital process is broken and displaced dorsally along the medial crack; the postorbital process is broken off, and when the original orientation of the supraorbital process is taken into account, it would have probably dorsally overlapped or contacted the zygomatic process ( Fig.17 View FIG ). Ŋe orbital margin of the supraorbital process is concave in dorsal view, and the dorsal surface of the supraorbital process is slightly anteroposteriorly arched. Ŋe posterior margin of the supraorbital process of the frontal is oriented transversely. Medially, the frontal drops sub-vertically from the vertex to the near-horizontal supraorbital process, which is horizontally oriented. Ŋe frontal drops near vertically from the vertex, so that it faces dorsolaterally, and the overlapping sheet of the parietal can be seen in dorsal aspect. Ŋe optic canal opens anterolaterally and is positioned posteriorly on the ventral surface of the supraorbital process of the frontal ( Fig. 12 View FIG ), defined by two ridges – the posteriorly placed infratemporal crest, and an anterolaterally directed ridge anterior to the canal. Ŋis anterior ridge is laterally confluent with the preorbital process, which projects slightly laterally.
Parietal
Ŋe parietal forms most of the medial wall of the temporal fossa, is exposed along the anterior portion of the vertex, and forms the ventral contribution to the nuchal crests on either side of the occipital shield ( Fig. 13B View FIG ). Ŋe lateral surface of the parietal in the temporal fossa is concave. Slightly anteromedial to the junction of the nuchal crest and the supramastoid crest of the squamosal, the parietal-squamosal suture descends ventrally along a bulge in the wall of the temporal fossa; it is oriented ventromedially, and does not turn posteriorly to form a long lateral finger of the parietal as in B. acutorostrata .
ONTOGENY
Several features of the skull suggest that the holotype cranium of B. bertae n. sp. represents an immature individual. Ŋese include relatively large occipital condyles, large petrosal fossae, and a relatively short compound posterior process of the petrotympanic. Additionally, several cranial sutures remain unfused, including the squamosal-parietal suture and the frontal-parietal suture; young ontogenetic age may explain the ease at which basicranial elements were lost or fractured (see Taphonomy). Ŋese features suggest that UCMP 219078 was not finished growing, and highlights the possibility that the morphology may not be representative of the adult form. In concert with the size of UCMP 219078 and development of these features in comparison with juveniles of extant Balaenoptera , the holotype of B. bertae n. sp. is interpreted as an old juvenile or subadult. Ŋe primary features used for diagnosing B. bertae n. sp. include the narrow transverse width of the vertex and the dorsolaterally facing surface lateral to the vertex, permitting the anterior wing of the parietal (and proximal portion of frontal) to be visible in dorsal aspect. Examination of juvenile and neonatal specimens of B. acutorostrata and B. physalus (CAS, USNM collections) indicates these features do not change during postnatal development. For example, small juveniles of B. acutorostrata (e.g., CAS 22180) exhibit a relatively wide apex of the occipital shield, and a vertical slope adjacent to the vertex.Ŋese diagnostic features can thus reasonably be interpreted as present in fully grown adults of B. bertae n. sp. Although the compound posterior process of the petrotympanic is relatively short, it is inflated with a blunt distal apex; in extant adult Balaenoptera spp., the distal apex is tapered. Examination of juvenile Balaenoptera indicates the distal apex is also tapered, unlike the condition in Balaenoptera bertae n. sp., suggesting an additional derived feature that does not appear to be affected by ontogenetic patterns observed in extant relatives. Furthermore, the posterior process extends laterally to the level of the lateral extremity of the paroccipital process, unlike immature specimens of extant Balaenoptera ; during ontogeny, the posterior process increases in length and lags behind the exoccipital during early development ( Bisconti 2001). Examination of the petrosal fossae of juvenile B. acutorostrata indicates that even in neonates, the fossae are smaller relative to skull size than in UCMP 219078, suggesting that the large size is autapomorphic for B. bertae n. sp., and possibly a paedomorphic trait. In summary, although several features suggest immaturity, other features suggest that growth was nearly completed; additionally, most of the diagnostic features for B. bertae n. sp. do not appear to be affected by ontogenetic changes in extant Balaenoptera .
TAPHONOMY
Ŋe skull of B. bertae n. sp. exhibits several noteworthy preservational features ( Fig. 16 View FIG ). UCMP 219078 was preserved in a monotonous succession of massively bedded, pervasively bioturbated finegrained sandstone. Ŋis facies, typically lacking any primary sedimentary structures due to thorough homogenization of the sediment by bioturbators, was interpreted by Norris (1986) as reflecting a middle shelf (or transition zone) depositional setting. Other evidence for a lag deposit such as abundant, large terrigenous or phosphatic clasts or abundant mollusks is absent from this stratum. Ŋe rostral bones (premaxillae, maxillae, and nasals), tympanic bullae, middle ear ossicles, and bodies of the pet- rosals are missing, and no postcrania were found during excavation. Ŋe vomer and other basicranial elements are shattered, and the left supraorbital process of the frontal and vomer are broken and rotated dorsally. UCMP 219078 was found ventralup in massively bedded, bioturbated sandstone with dispersed mollusks and pebbles. Ŋe cranium was likely too large to be overturned by currents and probably represents the original orientation of the skull upon arrival at the sea floor. Ŋe lack of postcrania suggests that the skull arrived on the sea floor as an isolated element. Ŋe lack of evidence for a lag deposit or erosional surface at this horizon suggests that postcrania were not dissociated by transport and submarine erosion.
Many cetacean carcasses float ventral up ( Schäfer 1972) and it is possible that the orientation of the skull reflects this; it also may reflect the hydrodynamically stable orientation of the skull as it sank. Ŋe rostral elements could have been lost during the bloat and float stage ( Fig. 16 View FIG A-C). Due to exaggerated movement of craniovertebral joint in modern floating cetacean carcasses, the skull is lost not long after the mandibles (e.g., Fig. 16E View FIG ; Schäfer 1972), which is reflected in some cetacean assemblages ( Bianucci et al. 2010). How the basicranial elements shattered is less clear, but some features of the fossil help to constrain the timing of deformation. Parts of the skull inside the concretion are shattered, indicating that this happened prior to formation of the concretion. Rotation and displacement of the vomer and supraorbital process of the frontal would not have been possible once entombed in sediment, and must have occurred prior to burial. Ŋe sequence of events for the holotype individual of B. bertae n. sp. is inferred as follows: bloating and floating of the whale after death (whether immediate or after refloating; Fig. 16A View FIG ); initial decomposition and loss of the jaws and rostral elements ( Fig. 16B, C View FIG ); advanced decay and skeletonization of the head results in the loss of the earbones ( Fig. 16D View FIG ), and eventual loss of the skull ( Fig. 16E View FIG ). At that time, the skull sinks to the sea floor ventral-up ( Fig. 16F View FIG ), and at some point between its arrival on the seafloor and burial, the vomer and supraorbital process rotate and come into contact with the seafloor ( Fig. 16G View FIG ). Eventually the skull is buried ( Fig. 16H View FIG ). Long after burial, diagenetic compaction fractures the basicranial elements ( Fig. 16I View FIG ), and a concretion forms around the braincase ( Fig. 16J View FIG ).
PHYLOGENETIC ANALYSIS
Balaenoptera bertae n. sp. was coded for the 150 characters of Marx (2011) as follows:
??????????????1???????????00012?210?1? ??11???01????????1012011130210210011?0 101?111?????????21??????????????????? ?????????????????????????????????????
Incorporation of B. bertae n. sp. into the matrix of Marx (2011) and cladistic analysis supported inclusion (with low branch support) of this new taxon within Balaenoptera ( Fig. 18 View FIG ). In the strict consensus tree, B. bertae n. sp. appears within a basal polytomy with Balaenoptera physalus , B. edeni , B. acutorostrata , and B. bonaerensis . Within Balaenoptera , the Peruvian fossil B. siberi is sister to a B. omurai + B. musculus clade, forming the only resolved clade within Balaenoptera ; however, this relationship is weakly supported ( Fig. 18 View FIG ). Topology among other mysticete taxa did not change relative to the phylogenetic results of Marx (2011: fig. 3), although branch and bremer support values slightly decreased across Balaenopteroidea, and support for Eschrichtiidae slightly increased ( Fig. 18 View FIG ). Most notably, the moderately supported B. borealis - B. bonaerensis - B. acutorostrata clade ( Marx 2011: fig. 3) collapsed in this analysis, perhaps owing to the inclusion of the somewhat incomplete B. bertae n. sp. Increased resolution of balaenopterid phylogeny would be possible with the inclusion of additional balaenopterid taxa into this matrix (such as D. luctoretmurgo , C. capellinii , P. quarantellii , and “ B. ” cortesi “var.” portisi) and additional balaenopterid-specific characters, including petrotympanic characters recently discussed by Ekdale et al. (2011), and other skull features such as a squamosal crease, which in the current study was only identified in modern and fossil species of Balaenoptera .
BALAENOPTERA BERTAE N. SP. AS A BALAENOPTERID Balaenoptera bertae n. sp. exhibits several balaenopterid characteristics, including an abruptly depressed
supraorbital process of the frontal, an anteriorly thrusted occipital shield that laterally overhangs the temporal fossa, and telescoping of the rostral elements and supraoccipital such that the parietal is only exposed as a narrow transverse band at the vertex ( Figs 11-15 View FIG View FIG View FIG View FIG View FIG , 17 View FIG ; Zeigler et al. 1997; Deméré et al. 2005; Bisconti 2007a, b, 2010a; Bosselaers & Post 2010). Balaenoptera bertae n. sp. shares with all extant Balaenoptera spp. and M. novaeangliae a broadly triangular occipital shield that laterally overhangs much of the temporal fossa so that it obscures most of the parietal and medial wall of the temporal fossa in dorsal view ( Figs 11 View FIG , 17 View FIG ; Deméré et al. 2005).In many fossil balaenopterids, the crania are less extremely telescoped and the supraoccipital is not thrust as far anteriorly and overhangs less of the temporal fossa (e.g., Archaebalaenoptera Bisconti, 2007 , “ B. ” cortesi “var.” portisi, C. capellinii , Protororqualus Bisconti, 2007 ) or only overhangs the anterior portion of the temporal fossa so that the posterior half of the medial wall is visible in dorsal aspect ( P. baulinensis , “ M. ” hubachi, “ M. ” miocaena). Most species of Balaenoptera , including B. bertae n. sp., exhibit a slight convex bulge along the medial wall of the temporal fossa at the position of the squamosal-parietal suture; Megaptera Gray, 1846 lacks this bulge, and is thus the only balaenopterid where the inner wall of the temporal fossa is completely obscured in dorsal view. In B. musculus , the occipital shield is transversely narrow, and the medial wall is so bulged that nearly the entire medial wall is visible in dorsal aspect (interpreted here to be a reversal).
Ŋe morphology of the squamosal varies widely within balaenopterids. Most fossil balaenopterids (e.g., A. castriarquati , “ B. ” cortesi “var.” portisi, P. baulinensis , and P. cuvieri ) appear to exhibit an elongate, laterally divergent zygomatic process with well-defined supramastoid crest that run along most of the length of the zygomatic process, while lacking a squamosal crease ( Zeigler et al. 1997; Deméré et al. 2005; Bisconti 2007a, b). An elongate and laterally flaring zygomatic process is regarded as the primitive condition amongst balaenopteroids (Deméré et al. 2005), and among fossil and extant balaenopterids, characterize all taxa (including Megaptera ) except for D. luctoretmurgo and Balaenoptera spp., which have relatively short and anteriorly directed zygomatic process. B. musculus is exceptional amongst extant Balaenoptera in its elongate and laterally flaring zygomatic process.Ŋe most extreme condition within balaenopterids characterizes B. acutorostrata , B. bertae n. sp., and D. luctoretmurgo , all of which have short and nearly anteriorly oriented zygomatic process. Postglenoid process morphology also varies within the Balaenopteridae , and it is short and indistinct in basal balaenopterids (e.g., Archaebalaenoptera , Parabalaenoptera , Protororqualus ) and posteriorly elongate in Balaenoptera , Diunatans Bosselaers & Post, 2010 , and Megaptera . Ŋese latter taxa (including B. bertae n. sp.) also exhibit a posterolaterally directed postglenoid process. Ŋe early diverging balaenopterid “ B. ” cortesi “var.” portisi also exhibits posteriorly elongate postglenoid process, but it is posteriorly and not posterolaterally oriented. A squamosal crease is present in taxa with anteriorly oriented zygomatic processes. While a squamosal crease is absent in D. luctoretmurgo ( Bosselaers & Post 2010) and B. siberi , it is present in all extant species of Balaenoptera and B. bertae n. sp., and absent in M. novaeangliae ( Kellogg 1922; Deméré et al. 2005). Ŋis feature may be a possible synapomorphy of Balaenoptera .
Balaenoptera bertae n. sp. may be distinguished from all extant balaenopterids by the lack of a fingerlike posterior projection of the pterygoid hamulus ( Fig. 11B View FIG ). Ŋe condition of this character is unknown in many fossil taxa, either because the ventral side of the skull has not been prepared (e.g., Archaebalaenoptera , B. siberi ), is completely missing ( Cetotheriophanes Brandt, 1873 , Plesiobalaenoptera Bisconti, 2010 ), or is too damaged to tell (e.g., “ B. ” cortesi “var.” portisi, “ M. ” miocaena, Parabalaenoptera ). “ Megaptera ” hubachi also appears to lack a posteriorly directed finger of the pterygoid hamulus. Fingerlike pterygoid hamuli are widely distributed among modern and fossil mysticetes such as cetotheriids sensu stricto, cetotheres sensu lato (sensu Bouetel & Muizon 2006), and extant Eschrichtius robustus (Lilljeborg, 1861) .
AS A MEMBER OF BALAENOPTERA
In general, B. bertae n. sp. exhibits characteristics that are relatively derived in comparison to most fossil balaenopterids. In comparison to extant species of Balaenoptera , B. bertae n. sp. is characterized by a mosaic of primarily derived and a few plesiomorphic characters. Aside from the derived features outlined above (also see Differential diagnosis), plesiomorphic characteristics of B. bertae n. sp. include a transversely narrow vertex and supraoccipital apex, and a supraor -
† Eschrichtioides gastaldii Strobel, 1881
Eschrichtius robustus (Lilljeborg, 1861)
4/56
† Parabalaenoptera baulinensis Zeigler, Chan & Barnes, 1997 † Archaebalaenoptera castriarquati Bisconti, 2007
† Protororqualus cuvieri (Fischer, 1829)
†“ Megaptera ” hubachi Dathe, 1983
†“ Megaptera ” miocaena Kellogg, 1922
Megaptera novaeangliae Brisson, 1762
Balaenoptera acutorostrata Lacépède, 1804
† Balaenoptera bertae n. sp.
Balaenoptera bonaerensis Burmeister, 1867
Balaenoptera borealis Lesson, 1824
Balaenoptera edeni Anderson, 1879
Balaenoptera physalus Linneaus, 1758
† Balaenoptera siberi Pilleri “& Pilleri, 1989
Balaenoptera musculus Linneaus, 1758
Balaenoptera omurai Wada, Oishi & Yamada, 2003
bital process of the frontal which is separated from the vertex by a dorsolaterally oriented and steeply sloping surface (of the frontal and parietal), instead of a vertically oriented (and laterally facing) surface ( Fig. 11 View FIG ). While a narrower vertex is plesiomorphic (Deméré et al. 2005; Bisconti 2007b), most basal balaenopterids exhibit a vertically oriented anterior lateral wall of the braincase and an abruptly depressed supraorbital process of the frontal, and the condition of the supraorbital process in B. bertae n. sp. may not necessarily be archaic. Likewise, B. bertae n. sp. does exhibit an abruptly depressed supraorbital process of the frontal, although offset by an oblique, dorsolaterally oriented surface rather than a vertical surface as in all other balaenopterids ( Fig. 14A View FIG ) and may be autapomorphic. An additional feature of B. bertae n. sp. is the apparent anteroposterior shortness of the vertex (judging by the anterior tips of the frontals, which approximate the posterior margin of the bony nares). In most extant balaenopterids, the anteriormost point of the supraoccipital is relatively far posteriorly, near the posterior margin of the supraorbital process of the frontal (or posterior to a transverse line dividing the supraorbital process into anterior and posterior halves). Ŋis condition characterizes most fossil balaenopterids as well, including Archaebalaenoptera , B. siberi , “ B. ” cortesi “var.” portisi, C. capellinii , “ M. ” hubachi, Parabalaenoptera , and Protororqualus . In B. bertae n. sp., B.bonaerensis , “ M. ” miocaena, and M. novaeangliae , the anterior margin of the occipital shield is positioned within the anterior¼ of the medial part of the supraorbital process of the frontal. Ŋis feature indicates that B.bertae n. sp. exhibits the most telescoped occipital shield of any species of Balaeno- ptera. Only one fossil balaenopterid appears to have a more anteriorly thrusted occipital shield, the Early Late Miocene “ M. ” miocaena; in this fossil taxon, the supraoccipital is thrust anteriorly to the level of the preorbital process of the frontal. Ŋe apex of the occipital shield is roughly at the postorbital process in basal balaenopterids like Archaebalaenoptera and Protororqualus , indicating that the condition in B. bertae n. sp. is relatively derived even when compared with extant balaenopterids. However, this may be an illusion caused by the apparent anteroposterior shortness of the supraorbital process of the frontal( Fig. 17C View FIG ) of B. bertae n. sp. relative to other balaenopterids,which appears to be an autapomorphic feature.
As demonstrated in the phylogenetic analysis, Balaenoptera bertae n. sp. represents a fossil species of the extant taxon Balaenoptera . Fossil species attributed to Balaenoptera by Deméré et al. (2005) include Balaenoptera davidsonii Cope, 1872 (Pliocene San Diego Formation of California), B. siberi (uppermost Miocene of Peru), Balaenoptera borealina Van Beneden, 1880 (Pliocene of Belgium), Balaenoptera minutis Van Beneden, 1880 (formerly Burtinopsis Van Beneden, 1872 ; Pliocene of Belgium), Balaenoptera rostratella Van Beneden, 1880 (Pliocene of Belgium), and Balaenoptera sibbaldina Van Beneden, 1880 (Pliocene of Belgium). Because P. J. Van Beneden did not designate any holotypes and because all the syntype material of the Belgian Pliocene taxa appear to represent multiple individuals or chimaeras, or are not considered to be diagnostic enough to be designated as lectotypes, Bosselaers & Post (2010) proposed that all Belgian Pliocene balaenopterid taxa (as well as Megapteropsis robusta Van Beneden, 1872 and Plesiocetus garopii Van Beneden, 1859 ) should be considered nomina dubia. Deméré et al. (2005) referred a skull (San Diego Formation, Pliocene, California) to B. davidsonii , which was previously only known by mandibles ( Deméré 1986), although this specimen now appears to represent a different taxon (T. A. Deméré pers. comm. 2011). While B. siberi was placed within Balaenoptera in the phylogenetic analysis of Deméré et al. (2005) and Marx (2011), Bosselaers & Post (2010) did not consider B. siberi to be a member of the genus as it exhibited “basal features”. Features that distinguish it from extant Balaenoptera and B. bertae n. sp. in- clude relatively slender and widely flaring zygomatic process, a supramastoid crest that continues nearly to the zygomatic apex, a broadly rounded supraoccipital apex, lack of a squamosal crease, and lack of a well-defined separation of the paroccipital process and squamosal in dorsal aspect.On one hand, several other basal balaenopterids exhibit a derived and wide vertex like Balaenoptera (e.g., Archaebalaenoptera , Parabalaenoptera ), while exhibiting plesiomorphic squamosal morphology (slender and widely flaring zygomatic processes and no squamosal crease); perhaps the morphology of B. siberi is exactly what could be expected of an archaic species within Balaenoptera . However, a recent phylogenetic analysis by Bisconti (2010b) suggests that B. siberi may lie outside Balaenoptera , as a sister taxon of Diunatans . Ŋe generic placement of B. davidsonii and B. siberi requires additional fossil material and phylogenetic studies. Ŋis study documents a convincing example of an extinct species of Balaenoptera from the Pliocene, and additional records of extinct(or extant) members of this genus from the Pliocene may await discovery.
REMARKS
Previous molecular analyses have utilized fossil calibrations in an attempt to constrain the timing of molecular divergence of various mysticete clades. Recent studies have yielded dates for the balaenopterid-eschrichtiid clade in the Middle to Late Miocene (13.8 Ma; McGowen et al. 2009;13-12Ma, Dornburg et al. 2012). However, molecular divergence dating is problematic for this clade due to the phenomenon of Balaenoptera paraphyly (by inclusion of Megaptera and occasionally Eschrichtius Gray, 1864 within Balaenoptera ), which is frequently recovered in molecular analyses ( Rychel et al. 2004; McGowen et al. 2009; Dornburg et al. 2012; but see Nikaido et al. 2006, and Steeman et al. 2009), although Megaptera and Eschrichtius fall outside the genus Balaenoptera in nearly all morphology-based cladistic analyses ( Steeman 2007; Marx 2011). Further complications arise when considering the choice of fossil calibrations. Dornburg et al. (2012) used “ M. ” hubachi as a calibration for the B. physalus - M. novaeangliae clade; however, some morphology-based analyses have failed to demonstrate a close relationship between “ M. ” hubachi and extant M. novaeangliae , and it is considered not to belong to Megaptera (Deméré et al. 2005; Bisconti 2010b). Problems of Balaenoptera monophyly notwithstanding, perhaps more appropriate fossil calibrations could be found in fossil species of Balaenoptera , as previously reported fossil records of Megaptera have turned out to be archaic balaenopterids with superficial similarities to Megaptera (“ M.” hubachi; “ M.” miocaena) or fragmentary remains from the Pliocene ( Whitmore & Kaltenbach 2008). Both B. bertae n. sp. and B. siberi have been found to fall within Balaenoptera ( Marx 2011; this study; but see Bisconti 2010b) although further testing is necessary for both taxa. Regardless of molecular divergence dating, fossils such as B. bertae n. sp. and B. siberi demonstrate that “true” Balaenoptera had evolved by the Early Pliocene, and possibly by the latest Miocene.



| UCMP |
University of California Museum of Paleontology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Balaenoptera bertae n.
| Boessenecker, Robert W. 2013 |
Diunatans luctoretmurgo
| Bosselaers & Post 2010 |
D. luctoretmurgo
| Bosselaers & Post 2010 |
Plesiobalaenoptera quarantellii
| Bisconti 2010 |
Archaebalaenoptera castriarquati
| Bisconti 2007 |
Parabalaenoptera baulinensis
| Zeigler, Chan & Barnes 1997 |
P. baulinensis
| Zeigler, Chan & Barnes 1997 |
Parabalaenoptera
| Zeigler, Chan, & Barnes 1997 |
Parabalaenoptera
| Zeigler, Chan, & Barnes 1997 |
Parabalaenoptera baulinensis
| Zeigler, Chan & Barnes 1997 |
Megaptera novaeangliae
| Brisson 1762 |