
Omoadiphas cannula, Mccranie, James R. & Cruz, Gustavo A., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199545 |
|
DOI |
https://doi.org/10.5281/zenodo.6197130 |
|
persistent identifier |
https://treatment.plazi.org/id/03E787CF-FFAA-FFFC-FF3F-6D6A412AFC5F |
|
treatment provided by |
Plazi |
|
scientific name |
Omoadiphas cannula |
| status |
sp. nov. |
Omoadiphas cannula sp. nov.
Figures 1–2 View FIGURE 1 View FIGURE 2
Holotype. UNAH 1740 (Universidad Autónoma de Honduras, Tegucigalpa), an adult male from Montaña de Peña Blanca, Sierra de Agalta, 4 km N of Catacamas, 14°53’26”N, 85°53’46”W, 1250 m elev., departamento de Olancho, Honduras, collected 13 September 1989 by Gustavo A. Cruz.
Paratype. UNAH 1732, an adult female, from the same locality and with the same collecting data as the holotype.
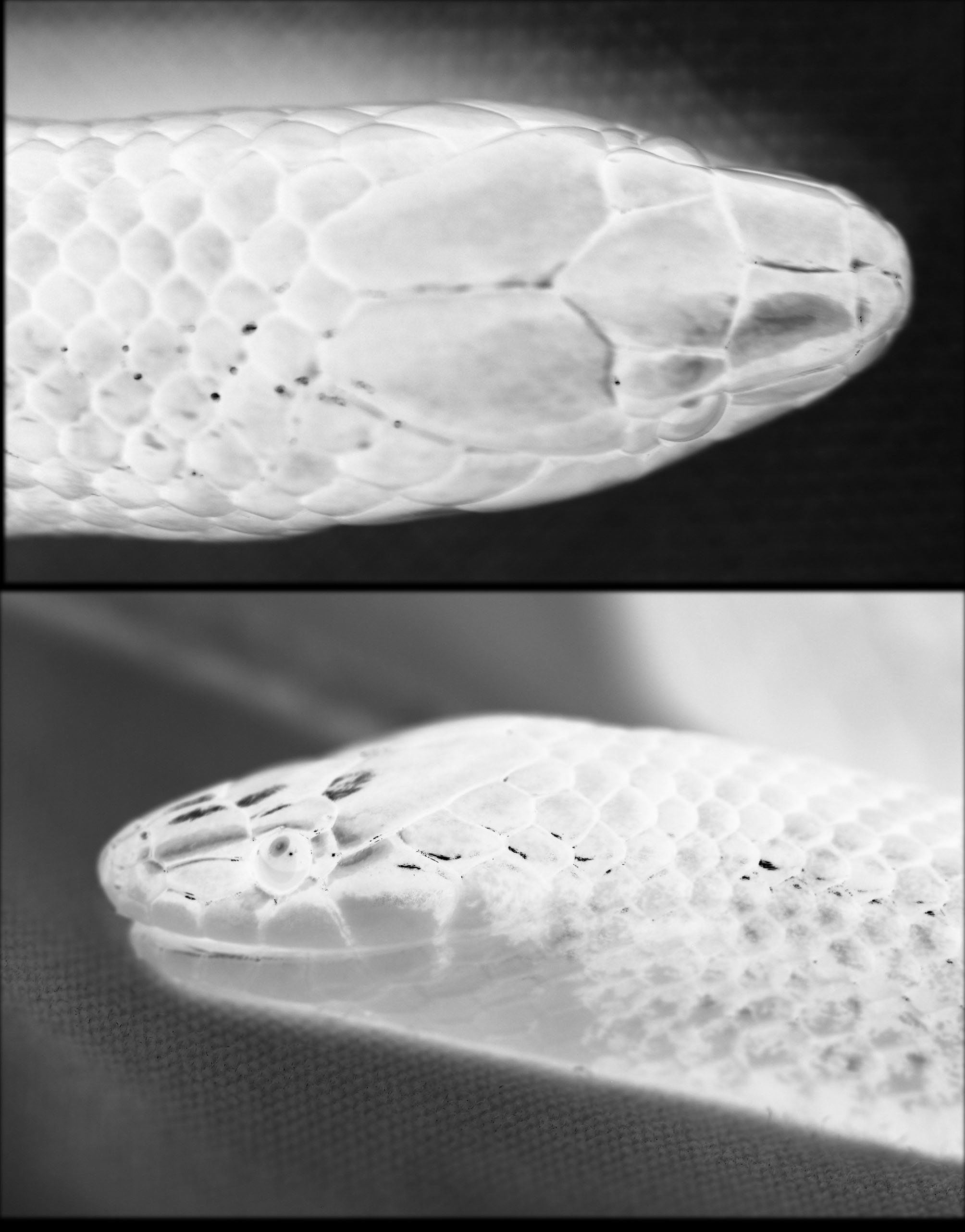
Diagnosis. Omoadiphas cannula can be distinguished O. aurula in having 47 subcaudal scales in the male (38–39 in O. aurula ; the “male” O. aurula with 35 subcaulals reported by Townsend et al. 2006 is in my opinion a female), six supralabials (seven), seven infralabials (eight), one postocular (two), the posterior nasal contacting the prefrontal (posterior nasal separated from prefrontal by loreal), a dorsal pattern of a dark stripe covering all of scale rows two and three (dark stripe on vertebral row and adjacent halves of scale rows two and three), and dark brown to nearly black ventral surfaces in preservative (pale yellow). Omoadiphas cannula differs from the single know specimen of O. texiguatensis (a subadult female) in having 31 subcaudals in the female (47 in O. texiguatensis ) and the dark dorsolateral stripe involving all of scale rows two and three on each side (confined to scale row three). The affinities of the three species of Omoadiphas appear to lie with a group of six other genera of snakes (see Köhler et al. 2001; McCranie & Castañeda 2004) that are part of a larger group referred to as “goo-eaters.” Omoadiphas cannula differs from the species of these six other genera in the following ways: from Adelphicos in having 17 dorsal scale rows (15), 154 ventral scales in the male and 167 in the female (117–155 in both sexes combined), and no anterior temporal (anterior temporal present); from all Atractus in having a divided cloacal scute (entire) and from select species of Atractus in lacking an anterior temporal (anterior temporal present in some Atractus ); from Chapinophis in having 154 ventral scales in the male and 167 in the female (178–196 both sexes combined), 47 subcaudal scales in the male (29–40 in both sexes combined), no anterior temporal (anterior temporal present), and no scale row reduction anteriorly on body (scale row reduction present); from Chersodromus in having 154 ventral scales in the male and 167 in the female (124–142 both sexes combined), 47 subcaudal scales in the male (maximum of 43), and a divided cloacal scute (entire); from all Geophis in having a divided cloacal scute (entire) and from select species of Geophis in lacking an anterior temporal (anterior temporal present in some Geophis ); and from Ninia in having smooth dorsal scales (strongly keeled), a divided cloacal scute (entire), a striped body pattern (stripes absent), and in lacking an anterior temporal (anterior temporal present).
Description of holotype. An adult male; TL 326 mm; SVL 266 mm; tail length 60 mm (18.4% of TL); HL 10.8 mm from front face of rostral to posterior end of mandible; HW 6.0 mm at broadest point (level of angle of mouth); head barely distinct from neck; snout rounded in dorsal view; eye length 1.1 mm; snout length 3.4 mm, about 3.1 times as long as eye; pupil subcircular; rostral about 1.7 times wider than high (1.2 mm x 0.7 mm); each internasal about as long as wide (0.6 mm x 0.7 mm); each prefrontal much larger than each internasal, about 1.3 times longer than wide (1.8 mm x 1.4 mm), bordering orbit above loreal and anterior to supraocular; median prefrontal suture (1.3 mm) 0.5 times as long as frontal; frontal broadly rounded anteriorly, strongly V-shaped posteriorly, about 1.3 times longer than wide (2.8 mm x 2.1 mm), slightly longer than distance from its anterior edge to tip of snout (2.6 mm); each parietal about 1.7 times longer than wide (4.1 mm x 2.4 mm), median suture (2.9 mm) about same length as frontal; parietals contacting seven nuchal scales; each supraocular longer than wide (1.6 mm x 0.7 mm), bordering orbit, contacting postocular, separated from loreal by prefrontal.
Nasal divided, anterior nasal contacting rostral, internasal, and first supralabial, posterior nasal contacting internasal, prefrontal, loreal, and first and second supralabials, nostril located in posterior portion of anterior nasal; loreal single, about 2.5 times longer than high (1.5 mm x 0.6 mm), lower edge contacting second and third supralabials, upper edge contacting prefrontal, loreal bordering orbit (no preocular); postocular single, about 1.7 times higher than long (0.5 mm x 0.3 mm); no anterior temporal, posterior temporal single, about 1.5 times longer than high (1.7 mm x 1.1 mm); supralabials 6–6, third and fourth bordering orbit, fourth and fifth contacting postocular, fifth also contacting parietal and posterior temporal, sixth contacting posterior temporal; mental about 2.5 times wider than long (1.0 mm x 0.4 mm), separated from chinshields by first pair of infralabials, which contact each other along ventral midline; chinshields about 2.4 times longer than wide (2.9 mm x 1.2 mm), not extending laterally to border of lip, separated from first ventral by two gular scales and four preventral scales; infralabials 7–7, first four contacting single pair of enlarged chinshields (their suture length 2.2 mm); a few tiny scale organs (tubercles) present dorsally and ventrally on head; dorsal scales in 17–17–17 transverse rows, smooth throughout, lacking apical pits and supracloacal tubercles; dorsal scales in 9 rows at level of tenth subcaudal; ventrals 154; cloacal scute divided; subcaudals 47, paired; tail spine pointed.
Maxillary teeth number 12–13 (right-left sides, respectively); longest teeth along middle portion of maxilla; teeth of middle portion of maxilla pointed, recurved, those of anterior and posterior portions blunter, less recurved; diastema absent; enlarged teeth absent posteriorly.
Color in alcohol: dorsal surfaces of head and body dark brown; slightly darker brown, broad longitudinal stripe involving all of scale rows two and three on each side of body; dorsal surface of tail slightly darker brown than that of body; supralabials creamy brown, except dark brown along upper edges; ventral surface of head creamy brown, that of body dark brown anteriorly, becoming progressively darker brown posteriorly; subcaudal surface darker brown than anterior two-thirds of body.
The hemipenes are only slightly everted, but agree with the description of the Omoadiphas aurula hemipenis in McCranie (2006) as follows: pedicel mostly nude, large naked pocket present on asulcate side; and truncus with one large basal hook on each side.
Variation in paratype. An adult female; TL 286 mm; SVL 255 mm; tail length 31 mm (10.8% of TL); ventrals 167; subcaudals 31. Otherwise the paratype agrees closely with the description of the holotype.
Distribution and habitat. Both specimens were collected under rotten logs during the late afternoon of 13 September, 1989 in slightly disturbed broadleaf forest (the Premontane Wet Forest formation of Holdridge, 1967). The type locality lies at 1250 m elevation, which at the time the specimens were collected was above the upper limits of the coffee farms on the slopes below. No other snake species were collected the day the type specimens were found, however, the first author returned to the type locality on 6 July 2010 and only found three other snake species ( Adelphicos quadrivirgattum , Ninia sebae , and Micrurus nigrocinctus ).
Etymology. The specific name cannula is formed from the Latin word canna (reed, cane) and Latin suffix – ulua (diminutive). The name refers to the Sierra de Agalta where this small snake was collected. The name Agalta is likely formed from the indigenous Pech language acatla (reed grass) and Tla (abundant).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |