
Diplura macrura ( C.L. Koch, 1841 )
|
publication ID |
https://doi.org/ 10.5852/ejt.2016.210 |
|
publication LSID |
lsid:zoobank.org:pub:E239C584-B41B-4AC7-89E1-E3D8A2EB921A |
|
DOI |
https://doi.org/10.5281/zenodo.5541656 |
|
persistent identifier |
https://treatment.plazi.org/id/03E7878E-4373-FFAC-BE59-E505FB8EFBA5 |
|
treatment provided by |
Plazi |
|
scientific name |
Diplura macrura ( C.L. Koch, 1841 ) |
| status |
|
Diplura macrura ( C.L. Koch, 1841) View in CoL
Figs 1–13 View Fig. 1 View Figs 2 – 13 , 26 View Fig. 26
Mygale macrura C.L. Koch, 1841: 38 , fig. 715 (♂).
Diplura bicolor Simon, 1889: 215 View in CoL (♀) syn. nov.
Thalerothele uniformis Mello-Leitão, 1923: 105 , fig. 4 (♂) syn. nov. Thalerothele minensis Mello-Leitão, 1926: 314 , figs 1-3 (♂) syn. nov. Thalerothele aurantiaca Mello-Leitão, 1943: 255 (♀) syn. nov.
Diplura bicolor View in CoL – Mello-Leitão 1937: 3, fig. 3 (♂). — Bücherl 1957: 385, figs 12, 12a. Diplura macrura View in CoL – C.L. Koch 1850: 75. — Raven 1985: 74, figs 18–23. Thalerothele uniformis – Bücherl, Timotheo & Lucas 1971: 128, figs 12–14 (syn. T. minensis , T. aurantiaca ).
Diplura uniformis View in CoL – Platnick 1993: 89 (transfer). — Silva-Moreira et al. 2010: 32. Linothele bicolor – Platnick 1998: 120 (transfer).
Diagnosis
Both sexes of this species have a strongly contrasting color pattern, with a reddish brown carapace and a dark brown abdomen, shared only with Diplura paraguayensis ( Gerschman & Schiapelli, 1942) . In mature D. macrura , the abdomen is uniformly dark brown, without spots or any visible markings. However, according to its original description ( Gerschman & Schiapelli 1942), D. paraguayensis has a blackish brown reticulate on abdomen dorsum. Moreover, the lyra of D. paraguayensis has 13 setae ( Gerschman & Schiapelli 1942: pl. ix; Schiapelli & Gerschman 1968, fig. 7), in contrast with the 7–9 setae in D. macrura . The bulb of D. macrura ( Figs 6–7 View Figs 2 – 13 ) has an embolic base much larger than in D. paraguayensis ( Schiapelli & Gerschman 1968: figs 14–15). Also, the embolus is about 2× longer than the bulb in D. macrura and 3× longer in D. paraguayensis . The spermathecae of D. macrura has a longer and thinner stalk ( Fig. 13 View Figs 2 – 13 ) than the females of D. paraguayensis from Argentina ( Goloboff 1982: 1). Also, the distal lobes are larger and spread over the distal third of the stalk in D. macrura , while they are smaller and concentrated in the apex in D. paraguayensis .
Type material
Mygale macrura : BRAZIL: ♂, holotype, Minas Gerais, São João del Rei, [1830], Sellow , Friedrich W. ( MfN 2083 ), examined .
Diplura bicolor : BRAZIL: 2 ♀♀, syntypes, Minas Gerais, Catas Altas, Caraça (coll. E. Simon, MNHN AR 4932 B337 ), examined (photos) .
Thalerothele uniformis : BRAZIL: ♂, holotype, São Paulo, E. of Garbe (MZSP?, MNRJ?), not located .
Thalerothele minensis : BRAZIL: ♂, holotype, Minas Gerais, Ouro Preto, Magalhães Gomes (Mello- Leitão collection 880, MNRJ 1360 ), not located .
Thalerothele aurantiaca : BRAZIL: ♀, holotype, Minas Gerais, Ouro Preto/Mariana: Itacolomi , O. Leonardos ( MNRJ 53945 ), not located .
Material examined
BRAZIL: 1 ♂, Minas Gerais, Parque Nacional Serra do Cipó , R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ) ; 1 ♀, 2 juvs, Caeté, 11–19 Feb. 1961, P. Vanzolini, H. Britski & N. Menezes leg. ( MZSP 6576 ) ; 1 ♂, Catas Altas, Reserva Particular de Patrimônio Natural Serra do Caraça , 24 Apr.– 1 May 2002, Equipe Biota ( IBSP 13148 ) ; 2 ♂♂, 1 juv., Lima Duarte, Parque Estadual de Ibitipoca , 21 Mar. 1997, A. Oliveira & B. Souza leg. ( IBSP 8390 ) ; 1 ♂, Parque Estadual de Ibitipoca , Apr. 1997, A. Oliveira & B. Souza leg. ( IBSP 8432 ) ; 1 ♀, juv., Ouro Preto, Estação Ecológica de Tripuí , R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4467 and MNRJ 4490 , respectively) ; 1 ♀, Estação Ecológica de Tripuí , 1220 m, 20°22'92.7" S, 43°33'29.3" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4502 ) GoogleMaps ; 1 ♂, Estação Ecológica de Tripuí , 1236 m, 20°22'93.2" S, 43°33'28.7" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4496 ) GoogleMaps ; 1 ♀ ( MNRJ 4487 ) , 1 ♂ ( MNRJ 4497 ) , 1 ♀ ( MNRJ), Estação Ecológica de Tripuí , 1246 m, 20°22'93.2" S, 43°33'28.7" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg GoogleMaps .; 1 ♀, Estação Ecológica de Tripuí , 20°22'6.62" S, 43°32'5.74" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4509 ) GoogleMaps ; 1 juv., Estação Ecológica de Tripuí , 20°22'9.70" S, 43°33'1.04" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4498 ) GoogleMaps ; 1 ♀, Estação Ecológica de Tripuí, 20°22'915" S, 43°33'227" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4489 ) ; 1 ♂, Estação Ecológica de Tripuí , 20°22'93.2" S, 43°33'26.7" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4505 ) GoogleMaps ; 1 ♂, Estação Ecológica de Tripuí , 20°22'93.2" S, 43°33'28.7" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4479 ) GoogleMaps ; 1 juv., Estação Ecológica de Tripuí , 20°22'93.2" S, 43°33'28.7" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ) GoogleMaps ; 1 juv., Estação Ecológica de Tripuí , 20°22'970" S, 43°33'104" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4492 ) ; 1 ♂, Estação Ecológica de Tripuí , 20°22'92,2" S, 43°33'13" W, 8 Jan. 2006, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ) GoogleMaps ; 1 ♀, Estação Ecológica de Tripuí , 20°22'93.2" S, 43°33'287" W, 7 Jan. 2005, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani M. leg. ( MNRJ 4333 ) ; Ouro Preto: 1 juv., Parque Estadual do Itacolomi , R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ) ; 1 juv., Parque Estadual do Itacolomi , R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4515 ) ; 1 ♂, Parque Estadual do Itacolomi , 1,306 m, 20°22'662" S, 43°32'574" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4495 ) ; 1 ♀, Parque Estadual do Itacolomi , 20°25'638" S, 43°30'399" W, R. Bertani, R. Martins, C.S. Fukushima & M.P. Pavani leg. ( MNRJ 4507 ) .
Description
Male (MNRJ 4496) ( Figs 3–9 View Figs 2 – 13 )
Carapace: 6.4 long, 5.0 wide. Abdomen: 6.6 long. Spinnerets: PMS 1.3 long; PLS, total length 6.8, basal article 2.0, middle 2.2, distal 2.6, respectively. Legs: see Table 1 View Table 1 . Carapace: length/width 1.3; flat, cephalic area slightly raised, thoracic furrows shallow and wide. Fovea: short, deep, recurved. Carapace covered with short, thin setae, interspersed with some longer and thicker setae; border with abundant long and thick setae pointing outwards, increasing in number towards posterior angles. Clypeus almost totally hidden by the bulging eye tubercle, frontal margin bearing 5 thick, long, erect setae. Eye tubercle: 0.6 long, 1.0 wide, area between posterior eyes covered with thin setae and bearing 4 thicker, longer setae. AME 0.3, almost spherical, but a bit longer than wide, set apart by 0.7× their diameter. ALE elliptical, much longer than wide, its length about 0.6× the AME diameter. PME small, with flattened lens, longer than wide, its length about 0.5× AME diameter. PLE elliptical, much longer than wide, its length a bit less than 0.7× AME diameter. PME and PLE clearly set apart by around 0.4× the PME length. Anterior eye row slightly recurved, posterior eye row recurved. Eye rows with similar width. Chelicerae: promargin with 11 teeth on left and 9 on right chelicera. Plectrum with 5 thick, long setae. Labium: length/width 0.8, no cuspules. Labio-sternal groove deep with elongated sigilla. Sternum: about 45% longer than wide, very similar to the sternum of D. lineata . Posterior angle in a blunt point, not separating coxae IV. Sigilla: three pairs, spherical, with a subtle increase in size from anterior to posterior, all near margin. Palp ( Fig. 5 View Figs 2 – 13 ): relatively short, without retrolateral spines, femur: d1–2–0, pl0–0–1, tibia pl 0–1–0, v1–2 –0. Tibia: length 2.3, width 0.9, short, incrassated, thinner at the basis and apex. Maxillae: length/width 1.5. Cuspules: 13 spread over ventral inner heel. Lyra at the ventral side of the maxilla, formed by 8 modified thick, long setae, increasing in size from basis to apex of the lyra, weakly curved at apical portion. Legs: Leg formula 4123. Legs covered with short, thin, horizontal black setae and with some longer, thicker, erect black setae. All tarsi with thin scopula, throughout the length of the article, divided by two series of thicker setae at the middle line of the ventral face. Metatarsus I with undivided thin scopula, covering the distal half of the retrolateral side. Metatarsus II similar to I, but with scopula on both sides. Metatarsi III–IV without scopula. All tarsi provided with numerous small cracks covering almost all the ventral and lateral faces, except by the basis and tip of the article. Leg I ( Figs 3–4 View Figs 2 – 13 ): tibia I with a relatively short distal retrolateral spur, curved and blunt, placed at its ventral corner. Megaspine pointed, slightly sinuous, especially on the apex, almost 2x longer than the spur. Metatarsus I with a distinct retrolateral tubercle placed ventrally at the beginning of the median third, conical, pointed and facing towards the tip of article. Ventrally, there is one spine near the apex and another one on the median third, placed much ahead of the tubercle. Fringe formed by many thick and long spiniform setae and spines (clasper) covering almost all prolateral side of the metatarsus ( Fig. 4 View Figs 2 – 13 ). Spines: leg I: femur d1–2–0, pld0–0–1, rld0–0–1, patella p0–0–1 left, p0 right, r0 left, 0–1–0 right, tibia p0–2–0, v0–1–1ap (apophysis), metatarsus p0–2–0, v0–1–1ap; leg II: femur d1–2–0, pld1–0–1 left, pld0–0–1 right, rld0–0–1 left, rld0 right, patella p0–1–1 left, p0–0–1 right, tibia p0–2–0, v1–1 –2ap left, v1 –0–2ap right, metatarsus p1–1–1 left, p1–1–0 right, v1–2 –2ap left, v1–2 –1ap; leg III: femur d1–2–0, pld0 left, pld0–1–1 right, rld0–1–1 left, rld0–2–1 right, patella p0–1–1 left, p0–1–1 right, r0–1–0 left, r1–1–0 right, tibia p0–2–0 left, p0–2–1 right, r1–1–1, v1–1 –2ap; metatarsus p2–2–1 left, p0–4–1 right, r1–1–1 left, r1–2–1 right, v0–4–3ap left, v0–3–3ap right; leg IV: femur d2–1–0 left, d1–2–0 right, rld0– 0–1 left, rld0–0–2 right, patella r0–1–0, tibia p0–1–0 left, p0–2–0 right, r1–2–1 left, r1–1–1 right, v2– 1 –2ap left, v1 –0–2ap right, metatarsus d1–0–0 left, d1–1–0 right, p1–2–1, r1–2–1, v1–5 –3ap. Claws: ITC without teeth. Teeth at STC: all claws with a small spur at the basis in both sides leg I: inner 6–7 and outer rows 5–6; leg II: inner row 9, outer row 8–9; leg III: inner row 6–7, outer row 6; leg IV: inner and outer rows 6–7. Bulb ( Figs 6–9 View Figs 2 – 13 ): piriform and conical in a retrolateral view, with length about ¾ of its width. Embolus about 2× longer than the bulb itself, with a wide base in relation to the globose part of the bulb and tapering towards the apex. Embolus clearly curved since its base and tapering from the basal third to the apex, clearly seen both on retrolateral and prolateral views. However, in dorsal view, the embolus is just slanted, almost straight. On ventral view, spermatic duct large at the base, tapering regularly towards the tip, but much thinner from the distal third on.
Female (MNRJ 4487) ( Figs 2, 10–13 View Figs 2 – 13 )
Total length 15.3. Carapace: 7.3 long, 5.8 wide. Abdomen: 8.0 long, 5.0 wide. Spinnerets: PMS 1.4 long; PLS, total length 5.9, basal article 2.3, middle 1.6, distal 2.0, respectively. Legs: see Table 2 View Table 2 . Females are very similar to males except by its bigger size and the following characteristics: carapace length/ width 1.3. Clypeus very narrow, with the frontal margin carrying five thick, long setae, facing forward. Eye tubercle with a thick seta on its anterior margin. Area between eyes with two longer and thicker setae. AME separated by around their diameter. PME and PLE less separated. Chelicera with 10–11 promarginal teeth, on the right and left chelicera, respectively. Plectrum with 6 thick, long setae. Maxilla with 16 (left) or 19 (right) cuspules. Lyra ( Figs 10–11 View Figs 2 – 13 ) as in male, formed by 7 modified setae. Tarsi I–II with scopula similar to male, but tarsus III with a very thin scopula and tarsus IV without scopula. Metatarsi I–II with undivided thin scopula, covering both sides and the ventral face. All tarsi provided with few small cracks covering only the median area of the ventral and lateral faces. Spines: leg I: femur d1–2–0, pld0–0–1; patella 0; tibia p0–0–1, v0–0–2ap (apophysis); metatarsus v0–3–2ap; leg II: femur d1–1–0, pld0–0–1; patella p0–0–1; tibia p0–0–1, v0–0–2ap; metatarsus v0–3–2ap; leg III: femur d1–0–0, rld0–0–2 left, rld0–0–1 right; patella p0–1–1 left, r0–1–0; tibia p1–1–0 left, p0–2–0 right, r0–2–0, v0–0–1ap left, v0–0–2ap right; metatarsus p1–2–1 left, p1–3–1 right, r1–1–1, v0–4–3ap; leg IV: femur d1–1–0 left, d1–2–0 right, rld0–0–1; patella r0–1–0; tibia p1–1–0, r2–1–0, v0–0–1ap left, v–0– 0–2ap right; metatarsus p0–2–1 left, p1–2–1 right, r2–1–2 left, r1–1–2 right, v1–3 –2ap. Spermathecae ( Fig. 13 View Figs 2 – 13 ): separated by about 70% of its length, with a thick stem, keeping a similar width up to the distal lobules. Stem curved forward from the median part on, bearing three large apical lobules of variable sizes.
Color pattern
Both sexes with carapace reddish brown, covered by abundant setae of a weakly coppery hue. Thoracic furrows and cephalic area darker. Eye area black. Legs, palps, chelicera and spinnerets dark brown, with a clear contrast to the color of carapace. Labium, sternum and leg coxae orange, with darker sigilla. Dorsum of the abdomen uniformly dark brown without markings or stripes, venter usually lighter colored than dorsum, but with a dark hue in some males.
Variation
Chelicera with 8–11 promarginal teeth. Lyra at the ventral side of the maxilla, formed by 7 to 9 modified setae ( Figs 10–12 View Figs 2 – 13 ). The maxillary cuspules may vary from 13 to 17 in males and from 16 to 19 in females.
Synonymy and notes
When describing Mygale macrura, C.L. Koch (1841: 39) indicated that the type-locality of the species was “WestIndien, St. Juan”. Nine years later, he transferred the species to his new genus Diplura , without mentioning the type-locality ( C.L. Koch 1850: 75). The type-locality indicated by Koch 1841 was cited differently by subsequent authors, for example, “Westindien” ( Ausserer 1871: 178), “St. Jean, Antilles” ( Simon 1903: 963), “West Indies” ( Petrunkevitch 1911: 60), or “Saint John, U.S. Virgin Islands” ( Muchmore 1993: 32). All those subsequent authors did not mention any additional specimen of D. macrura and were certainly restating the information given by C.L. Koch (1841).
Banks (1909: 155) was the first arachnologist to record D. macrura in Cuba, from Pinar Del Rio, as “ Ischnothele macrura Koch ”, but he made no comments on the implicit genus transfer, nor did he include any description of the specimens he examined. Lutz (1915: 77) and Petrunkevitch (1926: 27) also pointed out that D. macrura had previously been recorded from Cuba. While Lutz expressly cited Banks (1909), Petrunkevitch did not mention its source. However, he was probably also citing Banks (1909), as he stated that “ Diplura macrura C. Koch , has been also reported from Cuba”. In his paper on endemic spiders from Cuba, Alayón (2000: 38) also included D. macrura without any further comment. Currently, the only record for D. macrura included in the World Spider Catalog (2015) is Cuba, without any mention of Koch’s type-locality.
In her paper on Cuban spiders though, Bryant (1940: 260) considered Banks’ record a misidentification of Ischnothele longicauda Franganillo, 1936 . She based her opinion on the relative abundance of the latter species in Cuba. The family Dipluridae is scarcely represented in Cuba, with only I. longicauda and two species of Masteria L. Koch, 1873 recorded for the country ( World Spider Catalog 2015), not considering the erroneous D. macrura citation by Banks. Therefore, the only specimen of D. macrura undoubtedly cited for Cuba, West Indies or any other Caribbean locality is the holotype.
Surprisingly, an analysis of the holotype of D. macrura indicates that the type-locality was misinterpreted by Koch. The holotype label clearly indicates “Koch *. typ. S. João d. Rey. Sello” ( Fig. 1 View Fig. 1 , right). On the other hand, there is an additional label ( Fig. 1 View Fig. 1 , left) inside the vial that contains the type indicating “ Macrura N. Koch. * fg. 715. Cametá Sieber”, referring to Cametá, a locality in the state of Pará, northern Brazil. This additional label was probably wrongly added later in the vial, as there are no records for D. macrura from Pará. Up to now, all the specimens of D. macrura we examined came only from the state of Minas Gerais. Therefore, it is clear that the type-locality is really São João del Rei, central Minas Gerais state, Brazil. Friedrich Sellow (or Sello, 1789–1831), a famous Prussian naturalist, collected a large number of zoological specimens in Minas Gerais state, such as insects ( Papavero 1973) and birds ( Rego et al. 2013). Furthermore, most of Sellow´s specimens are also deposited at MfN.
Diplura bicolor was described from Caraça, Minas Gerais State, Brazil, based on two female syntypes ( Simon 1889: 215). It is, currently, placed in the genus Linothele , although it was considered a Diplura by former authors ( Mello-Leitão 1937; Bücherl 1957). This species was implicitly transferred to its current genus following Raven (1985: 182): “all alyrate Diplurinae […] are transferred to Linothele ”. As Simon (1889: 215) had not cited a lyra in his description, Diplura bicolor was regarded as a Linothele in all later spider catalogs (e.g., World Spider Catalog 2015). Notwithstanding the fact that the first description of a lyra in the maxilla of Dipluridae was made by Blackwall (1867), this structure was not mentioned again until the reevaluation of its form and function by Pocock (1896). So it is clear that Simon and other former authors had not dissected the maxilla of their specimens in search for a lyra. We have only been able to analyze photos of one syntype of Linothele bicolor ( Dipluridae Contributors, 2016 ), as it is currently in loan to another researcher, but the photos clearly show a lyra formed by 8 setae. Simon (1889: 216) clearly cited the characteristic contrasting color pattern found in D. macrura in the original description of D. bicolor . However, he also cited some light brown spots scattered on the posterior half of the abdomen. It is possible that the syntypes cited by Simon are immature or small mature females (15.3 mm long) since isolated light brown spots on the sides of the abdomen are usually found only in immatures of D. macrura , disappearing in older males and females. Besides the syntype photos, the examination of other specimens of D. bicolor from its type-locality (Caraça, Catas Altas) and nearby areas of Minas Gerais state allow the clear recognition of the species. The examined specimens agree very well to the original description of D. bicolor in color and size, except by the presence of a lyra. Furthermore, despite several collections on Caraça and other nearby localities, including extensive search for Mygalomorphae, we and other researchers had not been able to find any other Diplura or Linothele species from the same region. The comparison of the above cited specimens with D. macrura from central Minas Gerais state allowed us to establish this synonymy, based on the similar color pattern and lyra.
The male of D. bicolor was supposedly described by Mello-Leitão (1937: 3, fig. 3) and Bücherl (1957: 385, fig. 12–12a). However, both specimens clearly belong to other species. We were able to examine the “ allotypus ” of D. bicolor described by Mello-Leitão, from Gustavo da Silveira, Minas Gerais state (IBSP 3452). It is a male of an undetermined species of Nemesiidae , with a distinctive copulatory bulb and a very setose pedipalp ( Mello-Leitão 1937: fig. 3). Likewise, judging by the figures 12 and 12a by Bücherl 1957, the male he identified as D. bicolor is actually a Theraphosidae , with the typical two lobed tibial spur in leg I ( Bücherl 1957: fig. 12a) and a copulatory bulb also characteristic of that family ( Bücherl 1957: fig. 12).
Diplura uniformis ( Mello-Leitão, 1923) was originally described as Thalerothele uniformis by Mello- Leitão (1923: 105). He stated that the male holotype was collected in the state of São Paulo: São Paulo, by E. Garbe, and should be deposited in Museu de Zoologia de São Paulo (MZSP, old number 321). However, Bücherl et al. (1971: 119, 122) examined a male specimen from MNRJ, collected in Ouro Preto, labeled as type of T. uniformis by Mello-Leitão [MNRJ 192, MLPC 1056]. Since they were not able to find any type material of the species in MZSP, they considered the MNRJ specimen as the holotype of T. uniformis . They also stated that the type-locality and repository cited in the description were wrong ( Silva-Moreira et al. 2010: 32). Since a later examination of the MZSP collection by R. Baptista also did not produced any evidence of the holotype, we follow Bücherl et al. (1971) and Silva-Moreira et al. (2010) in considering the MNRJ specimen as the actual holotype of T. uniformis . The transfer of T. uniformis to Diplura was made by Platnick (1993) in his catalog, following Raven (1985).
Bücherl et al. (1971: 123) also considered T. minensis Mello-Leitão, 1926 and T. aurantiaca Mello- Leitão, 1943, both described from Ouro Preto ( Mello-Leitão 1926: 105 and 1943: 255, respectively) as synonyms of T. uniformis . Bücherl et al. (1971: 119) examined the female holotype of T. aurantiaca , collected in Itacolomi, Ouro Preto, by Othon Leonardos (MNRJ 53945), but not the male holotype of T minensis . In the description of Thalerothele minensis , it is stated that the holotype received the number MLPC 880. However, this number belongs to a type specimen of Opiliones, as Mello-Leitão wrote down in the catalogue of the part [or whole?] of his private collection purchased by MNRJ in 1929 ( Kury & Baptista 2004). Probably the wrong number in the original description was a misprint or a lapse ( Silva-Moreira et al. 2010: 32).
Unfortunately, we were not able to find any Diplura material from Ouro Preto in MNRJ collection, despite several attempts. Thus, the MNRJ specimen Bücherl et al. (1971) considered as the holotype of T. uniformis and the female holotype of T. aurantiaca had also been probably lost later on. We follow Bücherl et al. (1971) on the synonymies of T. minensis and T. aurantiaca with D. uniformis , considering that all species were described from Ouro Preto and the original descriptions do not allow the recognition of any diagnostic trait. T. minensis was diagnosed in relation to T. uniformis (= D. macrura ) by Mello- Leitão (1926: 11) based on the uniformly dark color of the abdomen, 7 setae on lyra and only 5 teeth on the promargin of chelicera. Nevertheless, the coloration of the venter in D. macrura males vary from a light hue to a dark color similar to dorsum, the number of setae on lyra varies between 7 to 9 and the number of cheliceral teeth varies from 8 to 11. The last character is a not reliable one, as it may be variable even in one specimen, as the 9 and 11 teeth on different sides of the male herein redescribed demonstrate. The illustrations of the holotype of T. minensis by Mello-Leitão (1926: figs 1–3) are poorly done and not diagnostic. The similarity in color, size and lyra structure indicates that it really is a synonym of D. macrura .
T. aurantiaca is just the female of T. uniformis (= D. macrura ), with the same color pattern and similar size as the males. Among the characters mentioned in the original description, only the 6 setae on lyra fall out of the range for D. macrura (7 to 9). However, the first seta in the lyra is smaller and thinner than the others ( Fig. 10 View Figs 2 – 13 ) and the number of setae increases over age. Therefore, it is probable that the 6 setae may represent a normal variation or indicate that the holotype was a small young female. The holotype vulva poorly illustrated by Bücherl et al. (1971: fig. 12) does not seem to be fully developed. Apart from the reasons mentioned above, only one species has been collected in Ouro Preto, in spite of many collecting trips resulting in abundant specimens of D. macrura . Notwithstanding the loss of the types of the three species from Ouro Preto, the synonymies also avoid the proliferation of nomina dubia. We compared specimens of Ouro Preto with specimens of D. macrura and this examination indicated that they all belong to the same species, as they share the same color pattern, structure of lyra and shape of male and female genitalia.
Habitat notes
D. macrura specimens have been found under fallen logs and rocks in relatively dry areas of Atlantic Forest or Cerrado vs Atlantic Forest ecotones. The spiders do not make funnel-webs, but applies silk to the ground or log cavities, sometimes building small entrance silk tubes.
Distribution ( Fig. 26 View Fig. 26 )
Known only from localities in central and south Minas Gerais state, Brazil.


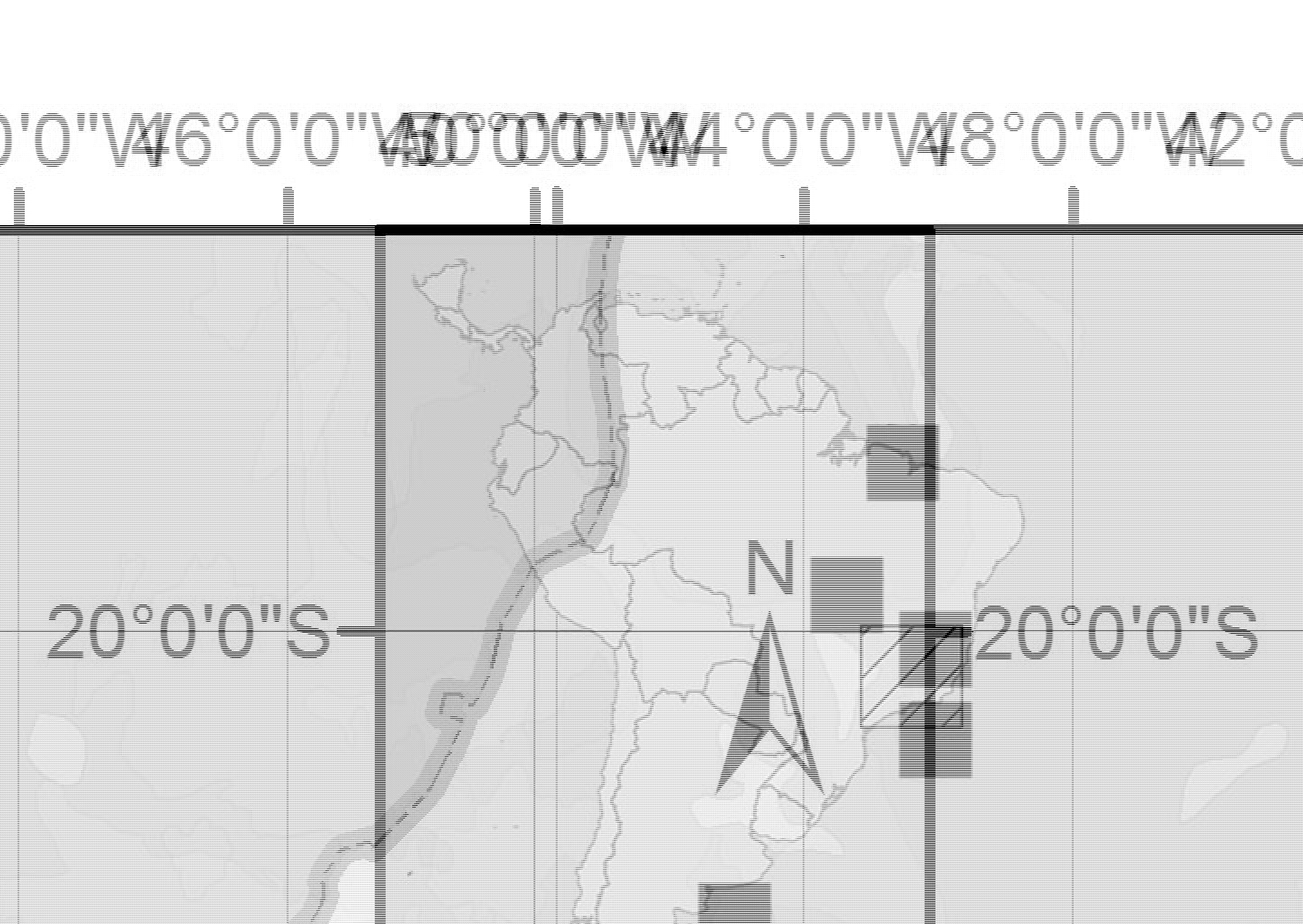
Table 1. Diplura macrura (C. L. Koch, 1841), ♂. Length of left leg articles (dorsal view).
| Leg I | Leg II | Leg III | Leg IV | |
|---|---|---|---|---|
| Fe | 5.5 | 5.1 | 4.7 | 5.9 |
| Pa | 3.2 | 2.9 | 2.4 | 2.5 |
| Ti | 4.2 | 3.8 | 3.2 | 4.4 |
| Mt | 4.0 | 4.1 | 4.4 | 6.2 |
| Ta | 3.3 | 3.2 | 3.1 | 3.6 |
| Total | 20.2 | 19.1 | 17.8 | 22.6 |
Table 2. Diplura macrura (C. L. Koch, 1841), ♀. Length of left leg articles (dorsal view).
| Leg I | Leg II | Leg III | Leg IV | |
|---|---|---|---|---|
| Fe | 5.6 | 4.8 | 4.4 | 5.2 |
| Pa | 3.4 | 3.0 | 2.6 | 2.9 |
| Ti | 3.6 | 3.3 | 3.0 | 4.2 |
| Mt | 4.0 | 3.5 | 4.2 | 5.6 |
| Ta | 3.0 | 2.8 | 2.8 | 3.0 |
| Total | 19.6 | 17.4 | 17.0 | 20.9 |
| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
| MNRJ |
Brazil, Rio de Janeiro, Sao Cristovao, Universidade do Rio Janeiro, Museu Nacional |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diplura macrura ( C.L. Koch, 1841 )
| Denis Rafael Pedroso, Pedro de Souza Castanheira & Renner Luiz Cerqueira Baptista 2016 |
Thalerothele aurantiaca Mello-Leitão, 1943 : 255
| Mello-Leitao 1943: 255 |
Thalerothele minensis Mello-Leitão, 1926 : 314
| Mello-Leitao 1926: 314 |
Thalerothele uniformis Mello-Leitão, 1923 : 105
| Mello-Leitao 1923: 105 |
Diplura bicolor
| Simon 1889: 215 |
Mygale macrura
| C.L. Koch 1841: 38 |