
Microplana hyalina Vila-Farré and Sluys, 2011
|
publication ID |
https://doi.org/10.1080/00222933.2010.536267 |
|
persistent identifier |
https://treatment.plazi.org/id/03E687FA-FFE6-BC1C-FE37-FB9CFD1BFD53 |
|
treatment provided by |
Felipe |
|
scientific name |
Microplana hyalina Vila-Farré and Sluys |
| status |
sp. nov. |
Microplana hyalina Vila-Farré and Sluys View in CoL , sp. nov.
( Figures 1 View Figure 1 , 6 View Figure 6 , Table 1)
Material examined
Holotype. ZMA V.Pl. 6858.1, Montsec , camí del Sant del Bosc, province of Lleida, Spain, 9 January 2002, sagittal sections on three slides.
Other material. ZMA V.Pl. 6859.1, Montsec , camí del Sant del Bosc, province of Lleida, Spain, 11 May 2008, sagittal sections on one slide .
Etymology
The specific epithet is derived from the Greek adjective hyalinos, like glass, and alludes to the hyaline colouration of the body.
Diagnosis
A small, hyaline Microplana species up to 5 mm long, with spherical penis bulb provided with a strong musculature and a distinct bulbar lumen; short and vertically oriented penis papilla; atrium divided into a cup-shaped cavity and a tubular distal
cavity; bursal canal with a distinctive, initially posteroventrally oriented loop-like curvature communicating with the posterolateral section of the copulatory bursa.
Description
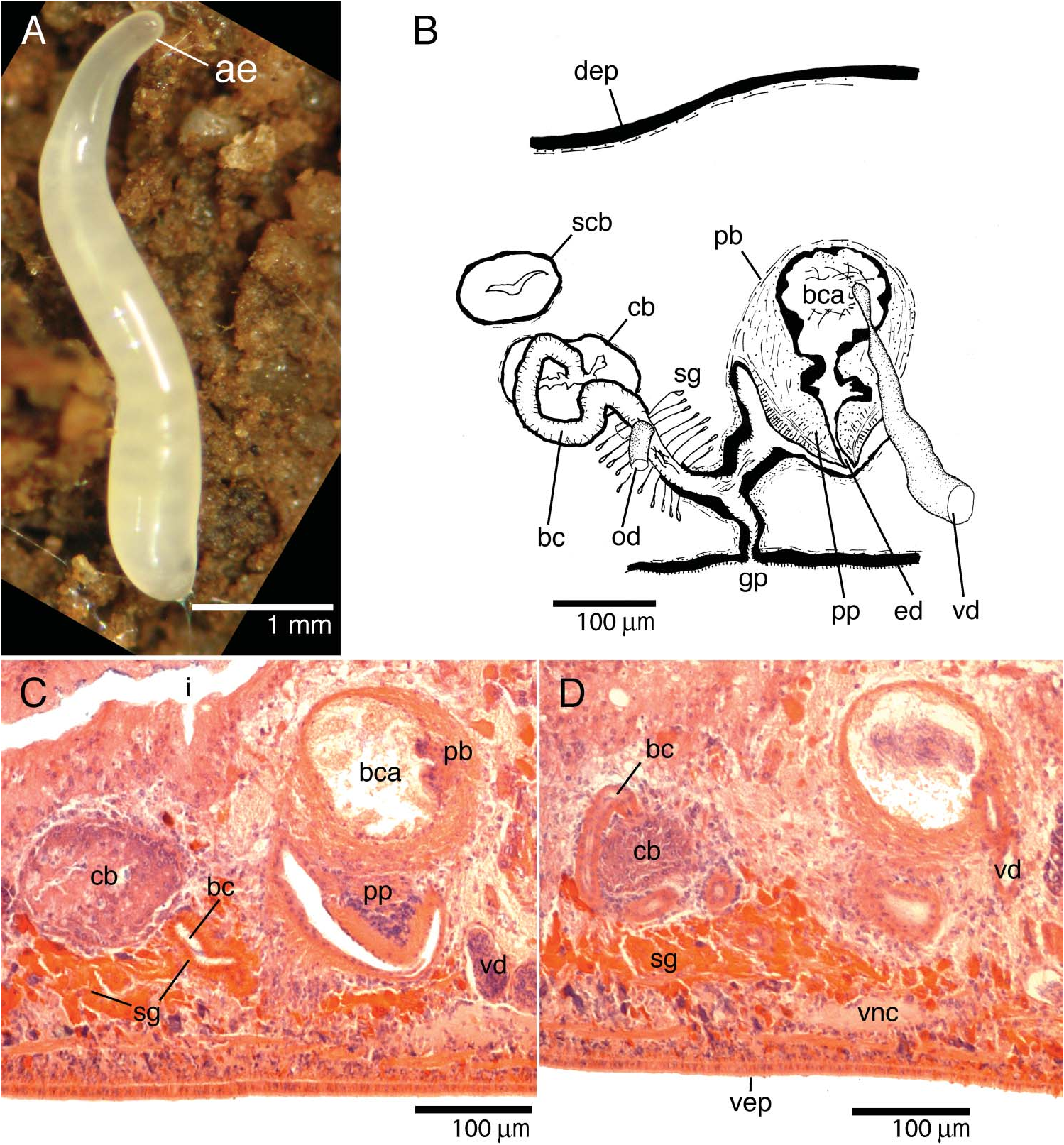
Living, sexually mature specimen ZMA V.Pl. 6858.1 about 5 mm long and about 0.5 mm wide, in elongated state ( Figure 6A View Figure 6 ). Cylindrical body tapers anteriorly to bluntly rounded point; posterior end also bluntly pointed. Dorsal and ventral body surfaces hyaline. Colouration and tiny size of preserved specimens prevented adequate observation of creeping sole. Two black eyes (eye cup diameter 19 µm in sections) a short distance anterior to brain.
Subepidermal longitudinal fibres of body musculature weak. In ventral body region numerous longitudinal fibres distributed in two mesenchymal bands, especially strong over and under ventral nerve cords. Dorsal longitudinal mesenchymal fibres run over intestine.
Short, cylindrical pharynx about one-eighth of body length, situated in posterior half of animal, in almost horizontal position. Outer epithelium of the pharynx ciliated, underlain by thin layer of longitudinal muscles, followed by thicker layer of circular muscles. Very thick outer layer of circular muscles underneath inner pharynx epithelium, intermingled with inner layer of longitudinal muscles fibres. In specimen ZMA V.Pl. 6859.1 posterior gut trunks meet at posterior end of body.
Mouth opening located at about one-third of distance between posterior wall of pharyngeal pouch and root of pharynx. In holotype mouth situated 2.1 mm from anterior end of body and 0.46 mm from gonopore.
About 12 ellipsoidal testes on each side of body. Follicles situated ventrally, occupying approximately one-quarter of dorsoventral diameter of body. Testes arranged in longitudinal rows, extending from some distance behind ovaries almost to root of pharynx.
Vasa deferentia narrow at level of penis before entering penis bulb, opening separately into lateral part of bulbar cavity ( Figure 6B View Figure 6 ). Strongly muscular, spherical penis bulb consists of intermingled longitudinal and circular muscle fibres. Bulb houses spherical and very spacious bulbar lumen ( Figure 6C View Figure 6 ) that narrows considerably before communicating with ejaculatory duct. Bulbar lumen and ejaculatory duct lined with nucleated epithelium underlain with layer of circular muscle fibres.
Vertically oriented penis papilla short and conical. Papilla covered with a thin, nucleated epithelium underlain with very thick, subepithelial layer of circular muscles, bounded by thick layer of longitudinal fibres. Male atrium consists of a dorsal cupshaped cavity and a lateral tubular part with diagonal orientation. Lining epithelium of atrium underlain with subepithelial circular muscle layer, thickened at tubular part, followed by thin layer of longitudinal muscles.
Ovaries situated above ventral nerve cords, occupying about one-quarter of dorsoventral diameter of the body, positioned between one-quarter and one-sixth of distance between brain and root of pharynx. Oviducts arise from ventral side of ovaries, where spermatozoa are observed. Oviducts run backwards laterally to ventral nerve cords; ducts turn dorsally and open separately into bursal canal.
Copulatory bursa spherical, lined with layer of tall vacuolated cells; not connected with gut. One specimen, ZMA V.Pl. 6859.1, had in its copulatory bursa remnants of an irregular structure (diameter 26 µm), presumably of a sclerotic nature. Origin of this structure not found in penis or atrial glands, but colour and texture suggest a sclerotic substance, suggesting that it may be part of a spermatophore. Sperm also present in this bursa. Secondary bursa-like structure not connected to the copulatory bursa nor to copulatory apparatus ( Figure 6B View Figure 6 ) present in holotype. Shell glands open into bursal canal mostly ventrally to openings of oviducts; bursal canal lined with nucleated and ciliated cells and surrounded by subepithelial layer of circular muscles and some scattered longitudinal muscle fibres. Bursal canal runs obliquely, i.e. posterodorsally, towards copulatory bursa from point of communication with atrium. After openings of oviducts, canal runs lateral to bursa with distinctive, initially posteroventrally oriented, loop-like curvature ( Figure 6B,D View Figure 6 ) before communicating with posterolateral section of copulatory bursa.
Discussion
Among the approximately 23 species of native land planarians known from Europe, two of which are described as new in the present paper, M. hyalina stands apart from the other species by the unique anatomy of its bursal canal, as well as by a unique combination of its internal and external characters. A hyaline body colouration is also found in M. humicola and M. aixandrei Vila-Farré et al., 2008 . Microplanana humicola , however, has a greenish anterior end, in contrast to M. hyalina . Regarding anatomical features, M. humicola has two pairs of dorsal testes and a genito-intestinal duct but a copulatory bursa is absent (cf. Schneider 1935). In contrast, M. hyalina presents 12 pairs of ventral testes and a copulatory bursa, but lacks a genito-intestinal duct.
Microplana aixandrei , a species endemic to the far southern part of the Iberian Peninsula but probably present also in other areas, is similar in size and colouration to M. hyalina and also produces spermatophores. The former has two testes on each side of the body, in contrast to M. hyalina , which has 12 testes on each side. In M. aixandrei the bursal canal is an obliquely running structure that communicates with the anterior section of the irregularly sac-shaped copulatory bursa; a sphincter is present in the proximal section of the canal. In contrast, in M. hyalina the canal runs laterally to the bursa and also shows a distinctive, initially posteroventrally oriented, loop-like curvature before communicating with the posterolateral section of the oval-shaped copulatory bursa; a sphincter is absent. The outer epithelium of the pharynx is ciliated only at the posterior part of the pharynx in M. aixandrei but completely ciliated in M. hyalina .
In addition to M. aixandrei , a copulatory bursa devoid of any connection with the intestine occurs also in M. howesi (Scharff, 1900) , M. mahnerti Minelli, 1977 , M. styriaca (Freisling, 1935) and M. grazalemica Vila-Farré et al. 2008 . However, in these species the bursal canal communicates with the anterior section of the copulatory bursa, whereas in M. hyalina the bursal canal communicates with the copulatory bursa through its posterolateral section. The gross morphology of the copulatory apparatus and the external appearance in these species are also very different from M. hyalina .
A dorsal connection of the bursal canal with the copulatory bursa is also present in M. monacensis (Heinzel, 1928) . However, in M. monacensis a genito-intestinal duct is present, whereas it is absent in M. hyalina . In addition, the horizontal penis papilla of M. monacensis is elongated but is conical and vertically inserted in M. hyalina .
Among the nine native species of Microplana from Africa, Madagascar and neighbouring islands, none has the external colouration and anatomy of the copulatory bursa and bursal canal of M. hyalina .
Unfortunately, M. gebavoeltzkowi from Madagascar is known only from its external appearance, namely dirty yellow with a broad grey-brown median band, which contrasts with the body colouration of M. hyalina .
Only one Asian species, Microplana indica ( Chaurasia, 1985) , possesses a copulatory bursa without intestinal connections (cf. Chaurasia 1985). The creamy dorsal surface of this species is marked with black spots making up a dorsal band. In contrast, in M. hyalina the dorsal surface is uniformly hyaline. Regarding anatomical features, M. indica has post-pharyngeal testes and a cylindrical penis papilla, whereas M. hyalina has pre-pharyngeal testes and a conical penis papilla.
Microplana teres , also from Asia, is described on the basis of an immature specimen with a slender penis papilla, contrasting with the short and blunt papilla of M. hyalina . Furthermore, the external colouration pattern is also different in both species.
The internal anatomy of the Asian M. ruteocephala is unknown but its external appearance differs much from M. hyalina (see above).
Only one American species has a copulatory bursa that is not linked to the intestine, M. costaricensis (De Beauchamp, 1913) . This species has a very long cylindrical penis and a bursal canal that communicates with the ventro-anterior section of the bursa. In contrast, in M. hyalina the penis is conical and the bursal canal opens at the posterolateral section of the copulatory bursa.
Various kinds of secretions may be associated with the sperm (cf. Souza and Leal- Zanchet 2004) but only very rarely in land planarians is the ejaculate enveloped by a true spermatophore of a clear sclerotic nature ( Sluys 1989b and references therein; Winsor 1998). Microplana hyalina is only the third species of land planarian for which such a spermatophore is described ( Winsor 1998). It has been suggested that in other species of land planarian other types of secretion, which do not completely surround the spermatozoa, may still function as spermatophore, facilitating the transfer of spermatozoa ( Souza and Leal-Zanchet 2004).
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.