
Pseudopaludicola canga
|
publication ID |
https://doi.org/ 10.5281/zenodo.282591 |
|
DOI |
https://doi.org/10.5281/zenodo.6176626 |
|
persistent identifier |
https://treatment.plazi.org/id/03E6336C-8E74-AB00-FF55-39A1C17EF985 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudopaludicola canga |
| status |
|
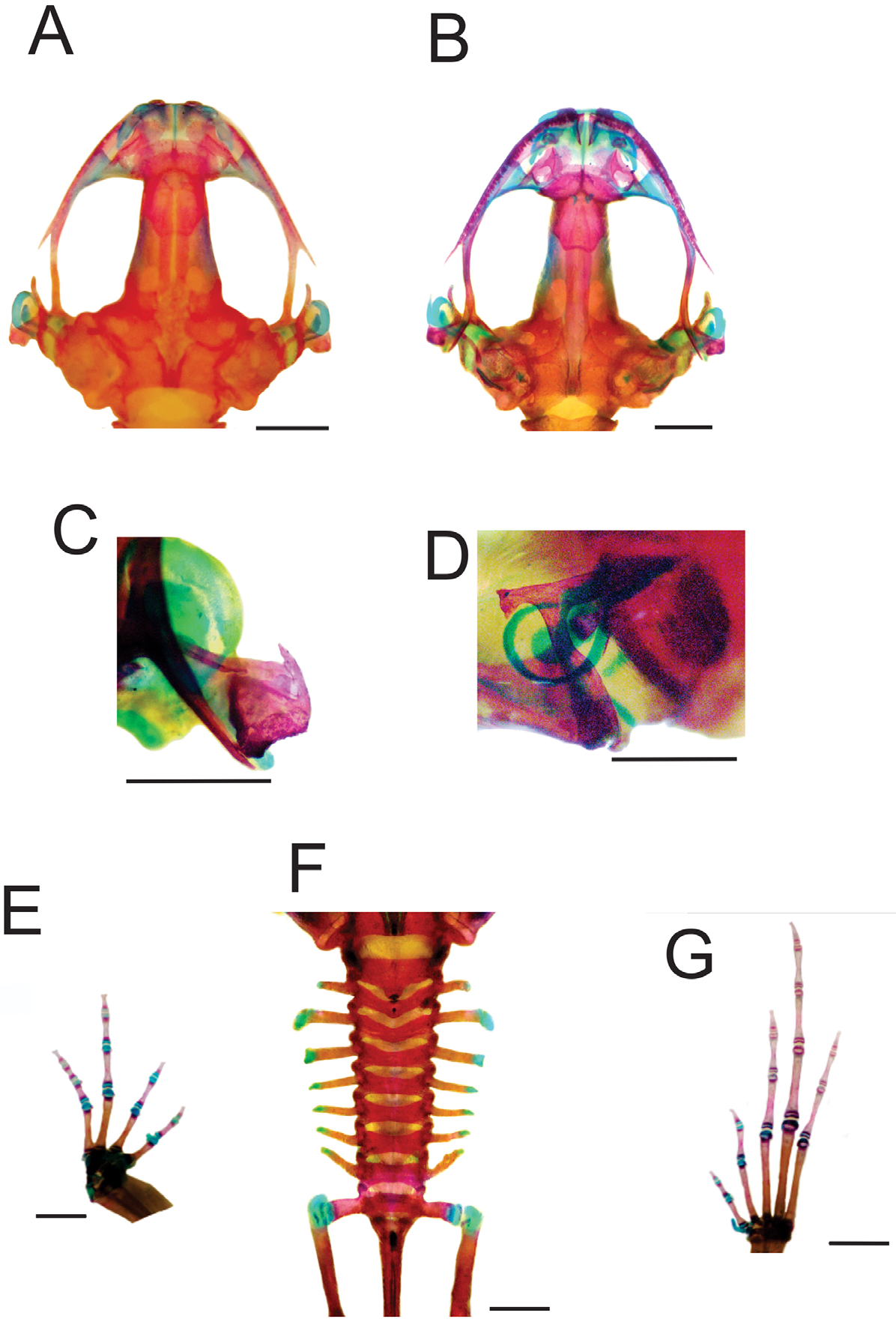
Pseudopaludicola canga osteology
( Figure 1 View FIGURE 1 )
Description based on MPEG 34368‒72 (three males; two females, Appendix I).
Skull wider than longer. Complete maxillary arch. Alary processes of premaxillae directed dorsally, parallel to each other (frontal view). Premaxilae bearing 7‒10 teeth and maxillae with 23‒28 curved and pedicellated teeth. Nasal narrow and ovoid, with irregular inner margin and posterior end pointed. The nasals are separated from each other and do not overlap the maxillary. In dorsal view, the nasal bone slightly overlaps the sphenethmoid (but does not overlap the sphenethmoid in MPEG 34368). Anterior margin of the sphenethmoid not reaching the middle of the choana, posterior margin W-shaped. In dorsal view, the sphenethmoid is short with a thin internasal septum. Frontoparietals with irregular inner margins that leave the frontoparietal fontanelle almost hidden ( Fig. 1 View FIGURE 1 A). Exooccipitals separated by a wide mineralized stripe. Optic foramen large sized, ovoid and placed on the junction between the prootics and the sphenethmoid (in MPEG 34368, the trochlear nerve exits from the neurocranium by an independent hole, placed anterodorsal to the optic foramen); Oculomotor foramen of narrow diameter, located posterior to the optic foramen (exit of cranial nerve III). Prootic foramen oval, placed posterior to the optic capsule (cranial nerve V). Jugular foramen with two holes (cranial nerves IX y X). Triradiate pterygoid. The posterior ramus of the pterigoid is longer than medium and is separated from the quadrate, while the medium is away from the wings of the parasphenoid and supported by cartilage on the otic region of the skull. Anterior ramus separated from the palatine. Quadratojugal with maxillary process reduced ( Fig. 1 View FIGURE 1 C). Triradiate parasphenoid with the tip of the cultriform process of irregular shape (rounded in MPEG 34368, 34370). Parasphenoid wings slightly laid back. Septomaxilary half-ring shaped (sickle-shaped in MPEG 34369, 34372). Neopalatines poorly developed, not overlapping the maxillae. Prevomers irregularly shaped, wider in middle region and without teeth. Squamosal with the ventral ramus wider at ventral end, zygomatic ramus shorter than otic with a descendent process welldeveloped ( Fig. 1 View FIGURE 1 D). Tympanic annuli cartilaginous incomplete, dorsally open with the posterior end parallel to the otic ramus of the squamosal. Plectrum with uniform width. Cartilaginous operculum, rounded.
Hyoid plate cartilaginous, mineralized at the center. Anteromedial processes short and divergent (parallel in MPEG 34370‒1). Postero- and anterolateral processes absent. Posteromedial processes short with cartilaginous ends.
The mandible has a thin dentary covering the lateral margins of the Meckels cartilage. The inner part of this cartilage is covered by the angular, which is thin and without coronoid process evident. The articular region of the mandible is cartilaginous and the dentary plain, without any serrate structures such as denticles.
The larynx has the arytenoid as two small oblong shells and the cricoid forming a complete ring. Esophageal process long and thin. Muscular processes thin. Cardiac processes poorly developed. In the females (MPEG 34370‒1), the cricoid is thinner and the arytenoids smaller.
Arciferal pectoral girdle, with the left epicoracoid cartilage slightly superimposing the right one. Clavicle thin, posterior margin concave, separated from the acromial region of the scapula by an anterior projection of the procoracoid. Scapula with pars acromialis and glenoidalis poorly developed. Anterior process of the suprascapula developed. Omosternum cartilaginous, without distal expansion (expanded in MPEG 34369‒70). Sternum simple, not bifurcated, with osseous mesosternum and cartilaginous xiphisternum expanded.
Humerus with deltoid crest developed. Radius-Ulna fused, leaving a groove between them, which begins at the union of the carpal elements and extends to two thirds of the length of both bones. Carpal composed by five elements: radial, ulnar, Y element, distal carpal V–IV–III and distal carpal II. Prepollex with osseous base and one mineralized long segment. Phalangeal formula: 2–2–3–3. Terminal phalanges knobbed, with central groove ( Fig. 1 View FIGURE 1 E).
Vertebral column with eight procoelous presacral non-imbricate vertebrae ( Fig. 1 View FIGURE 1 F). Cotilar facets of the atlas separated (Type I of Lynch 1971). The atlas body is the widest of the vertebrae. In dorsal view; the anterior margin of the atlas makes contact with the presacral II through a narrow neural process. Sacral diapophysis narrow, not expanded. Urostile with dorsal spine well-developed and exit of the spinal nerve X not visible. Urostilar articulation bicondylar. Anterior portion of the illium articulate with the ventral surface of the sacral diapophysis. Pubis cartilaginous. The hind limbs have the tibio-fibula fused at the distal end. Tarsal composed by: central, distal tarsal I, and distal tarsal II–III. Prehallux comprises an osseous base and one cartilaginous segment. Phalangeal formula: 2–2–3–4–3. Toe tips knobbed with central groove ( Fig. 1 View FIGURE 1 G).
Additionally, all the specimens examined present the same combination of sesamoid bones and osseous lateral projections:
At the anterior extremities the metacarpal epiphysis and some phalanges have small lateral projections. In the forelimbs, in dorsal view, there is a small rounded sesamoid bone over the distal carpal V–IV–III, in the tendon of the muscle extensor carpi ulnaris. In ventral view, the palmar sesamoid (embedded in the m. digitorum longus) has an irregular quadrangular shape. Also present are glide sesamoids on the flexor tendons of the hands, at the junction of the proximal and medial phalanges in all the fingers.
The hind limbs have lateral projections at the epiphysis of some metatarsals and phalanges and a constant sesamoid bones distribution: S. graciella (at the union between femur-tibia fibula, embedded at the tendon of the m. gracilis major); sesamoid cartilage (in the tendon of the m. plantaris profundus) and two plantar sesamoids with ovoid shape and similar size, placed at the tendons that form the aponeurosis plantaris.
Additionally, near the distal end of the sacral diapophyses there are a couple of sesamoids (S. of the sacral vertebrae) with elongated shape and similar width to the diapophyses.
Comments about toe tip digit IV. As previously mentioned by Lynch (1989), P. boliviana and P. ceratophyes have additional pads or cutaneous expansion at the end of the digits, reflected in the external morphology as wider T-shaped phalange ( Fig. 2 View FIGURE 2 E–F). On one hand, the T-shaped phalanges of P. l l a n e r a are the narrowest, but maintain a clear lateral expansion of the bone ( Fig. 2 View FIGURE 2 D), while in P. pusilla the toe tips are expanded forming a “robust T” ( Fig. 2 View FIGURE 2 C). In both cases, the width of the bone is not visible in the external morphology, so the end of the fingers is not expanded laterally (see Lynch 1971). However, all the species included in the P. pusilla group have the terminal phalanges T-shaped without a central groove, with the toe tips wider than the base of the last phalange. On the other hand, all the species unassigned to any group have the tip of the phalanges knobbed, with a central groove with the fingertips narrower than the base of the last phalange ( Lynch 1989; Lobo 1995), as is shown for P. falcipes ( Fig. 2 View FIGURE 2 B). In P. canga the toe tips are not T-shaped as mentioned by the original authors ( Fig. 2 View FIGURE 2 A) and this characteristic has important consequences for the diagnosis of the species.
Pseudopaludicola canga external morphology. Giaretta and Kokubum (2003) provided a detailed description of the holotype (ZUEC 6088). In our study we agreed with the previous description, adding some features of interest for the diagnosis of the species. We disagreed with the authors about the presence of dorsal folds. The holotype and remaining vouchers examined in this study have no dorsal folds. Additionally the vocal sac has a central fold (the vocal sac is more developed than other taxa (except P. serrana and P. murundu ), and in preservative forms a fold in the middle of the vocal sac). The toe tips in hands and feet are not expanded laterally, and are very similar to all species of Pseudopaludicola unassigned to any group. The absence of T-shaped phalanges is reflected in the fingertips being narrower than the base of the phalanges. Additionally, the inner thighs have a light brown background finely spotted with black.
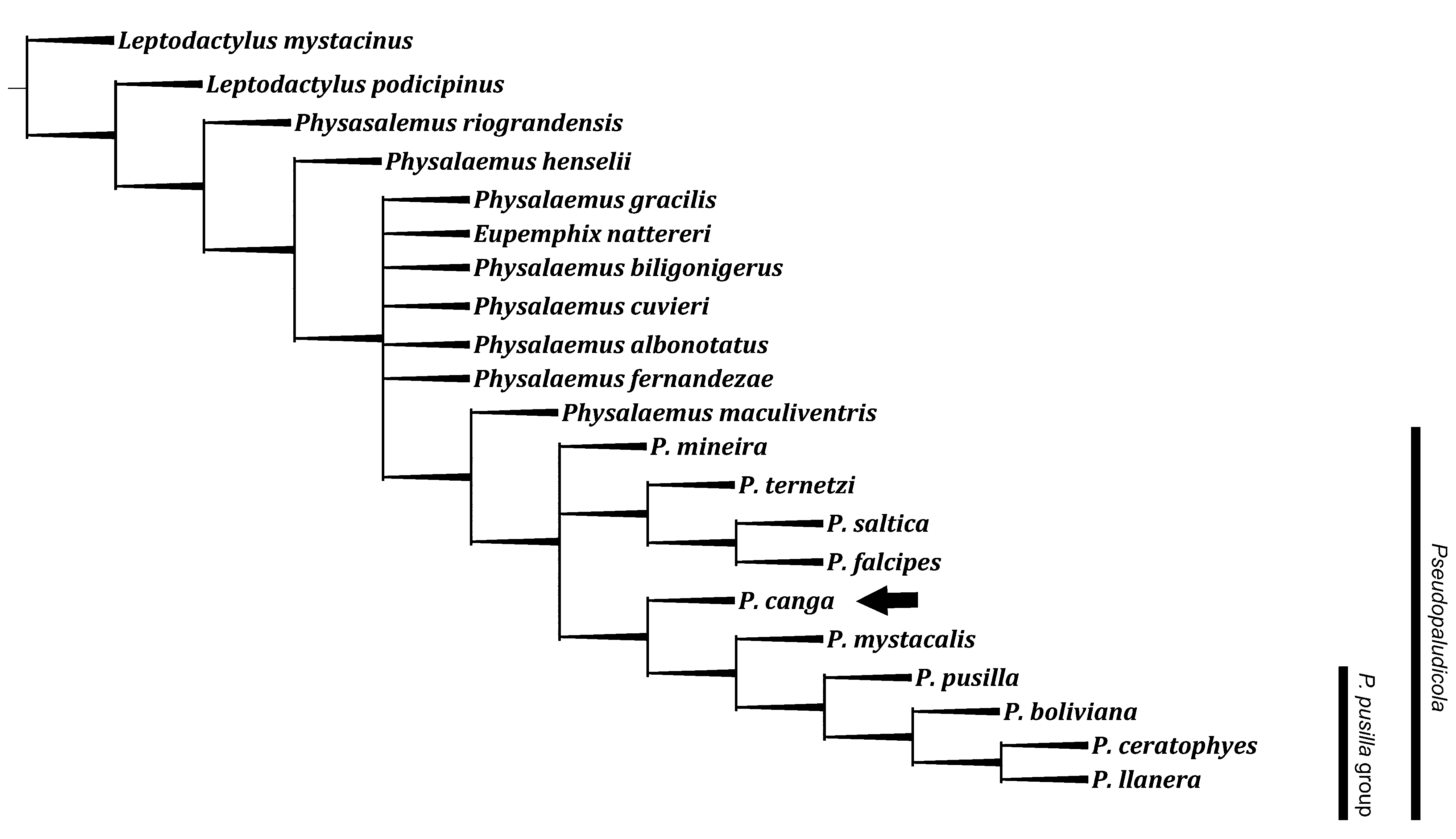
Taxonomy of P. c a n g a. The phylogenetic analysis obtained 16 maximum parsimony trees (length: 51 steps, consistency index 0.471; retention index 0.690). The strict consensus of this analysis is shown in Figure 3 View FIGURE 3 . Using the same character state and analysis methodology as Lobo (1995), P. canga has basal position in the clade formed by P. mystacalis (P. p u s i l l a ( P. boliviana (P. l l a n e r a + P. ceratophyes ))) in all most parsimonious trees as shown in the consensus tree ( Figure 3 View FIGURE 3 , black arrow). The unique monophyletic group recovered is the P. pusilla group, including P. pusilla ( P. boliviana (P. l l a n e r a + P. ceratophyes )), supported by the presence of T-shaped toe tips. The remaining species are unassigned to any group ( Fig. 3 View FIGURE 3 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.