
Unio tumidiformis Castro, 1885
|
publication ID |
https://doi.org/ 10.1080/00222930902993724 |
|
persistent identifier |
https://treatment.plazi.org/id/03E5EB5D-FFFC-490B-FDA5-FE77FDE5FCDF |
|
treatment provided by |
Felipe |
|
scientific name |
Unio tumidiformis Castro, 1885 |
| status |
|
Unio tumidiformis Castro, 1885 View in CoL
Synonyms
Unio batavus Morelet 1845: 109 View in CoL (not Unio crassus batavus Maton and Rackett, 1807 View in CoL ).
Unio sadoicus Castro, 1885: 284 (Sado river, Portugal; types lost); Locard 1899: 261; Azpeitia 1933: 249.
Unio macropygus Castro, 1885: 286 (Sado river, Portugal; types lost); Locard, 1899: 262; Azpeitia 1933: 251.
Unio eupygus Castro, 1885: 287 (Sado river, Portugal; types MNHN 20840 and Castro Collection n.° 47); Locard 1899: 262; Azpeitia 1933: 252.
Unio baeticus Kobelt, 1887: 55 View in CoL , fig. 496 (Guadaira river, Seville, Spain; types lost); Azpeitia 1933: 254, pl. IV, fig. 23.
Unio conimbricus Kobelt, 1893: 99 , pl. 180, 1133 (Mondego river, Pereira, Portugal; types lost).
Unio callipygus Drouët, 1893: 65 , pl. II, fig. 2 (Guadiana river, Ciudad Real, Spain; types lost); Azpeitia 1933: 243, pl. IV, fig. 21.
Unio jourdheuili Azpeitia 1933: 246 , pl. IV, fig. 22 (not Unio jourdheuili Ray in Locard, 1882).
Unio crassus batavus Haas 1940 View in CoL and 1969 (not Unio crassus batavus Maton and Rackett, 1807 View in CoL ).
Unio cf. crassus Reis 2006 View in CoL (not Unio crassus Philipsson, 1798 View in CoL ).
Type locality
Portugal, River Sado .
Type
The type series for U. tumidiformis is split between the Museu do Instituto Zoológico Augusto Nobre , Porto, Portugal (two specimens glued to cardboard, Castro collection no. 47) and the Muséum National d’Histoire Naturelle, Paris, France (21 specimens, MNHN 20839 About MNHN ). The Paris specimens are in the Locard collection, to whom Castro donated part of his material; Castro’s handwriting can still be identified in some shells and labels of this collection .
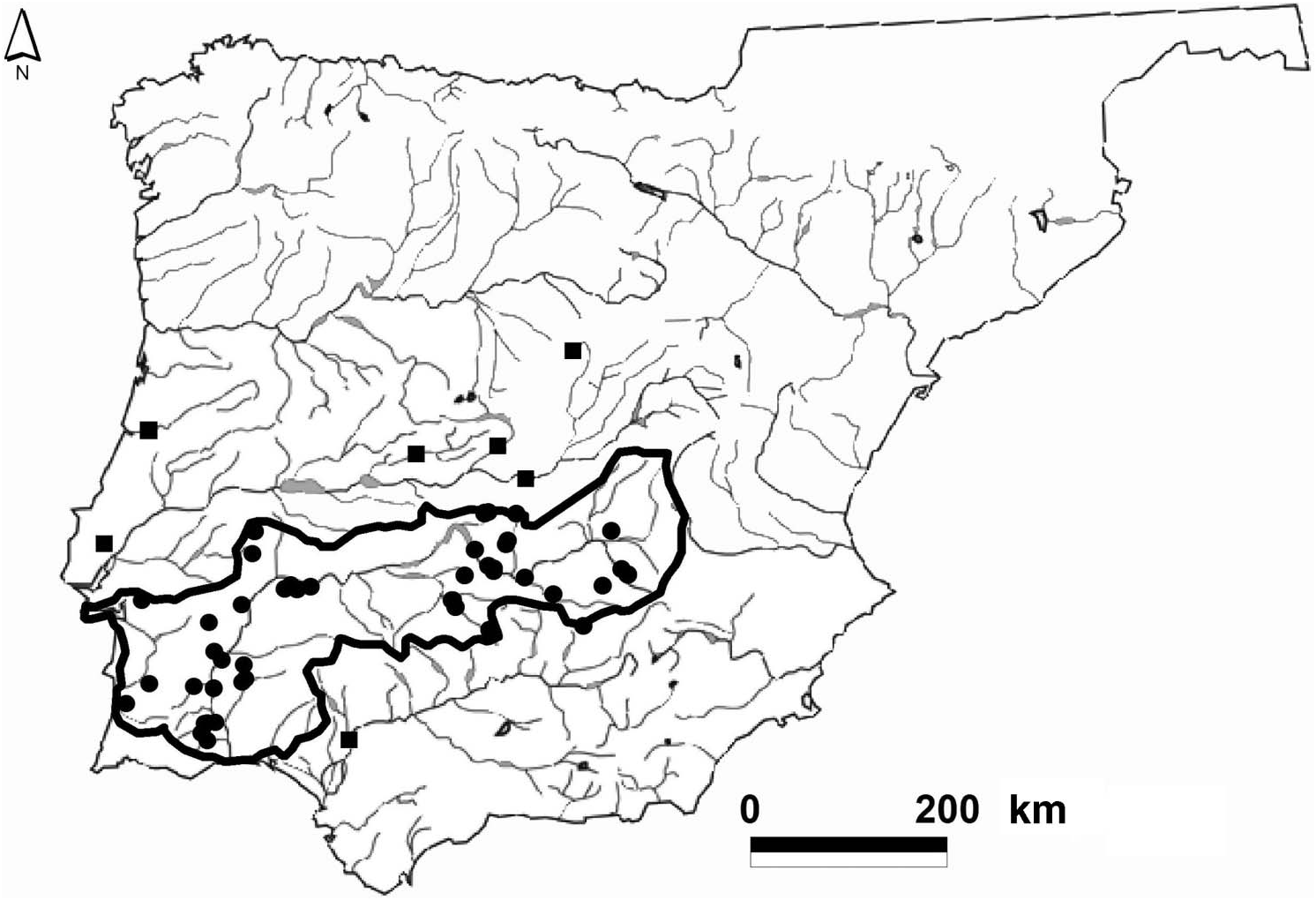
Distribution
South-western river basins of the Iberian Peninsula: Mira ( Portugal), Sado ( Portugal) and Guadiana ( Portugal and Spain) ( Figure 1 View Figure 1 and Table 1). Possible occurrence in
the Tagus and Guadalquivir basins ( Table 2). A single specimen recorded from the Mondego basin in Portugal ( Kobelt 1893; Table 2).
Habitat
Temporary Mediterranean-type streams. Sandy or muddy substrate of pool areas that do not dry up during the dry season. Habitat threatened by change of hydrological regimen of the streams.
External morphology
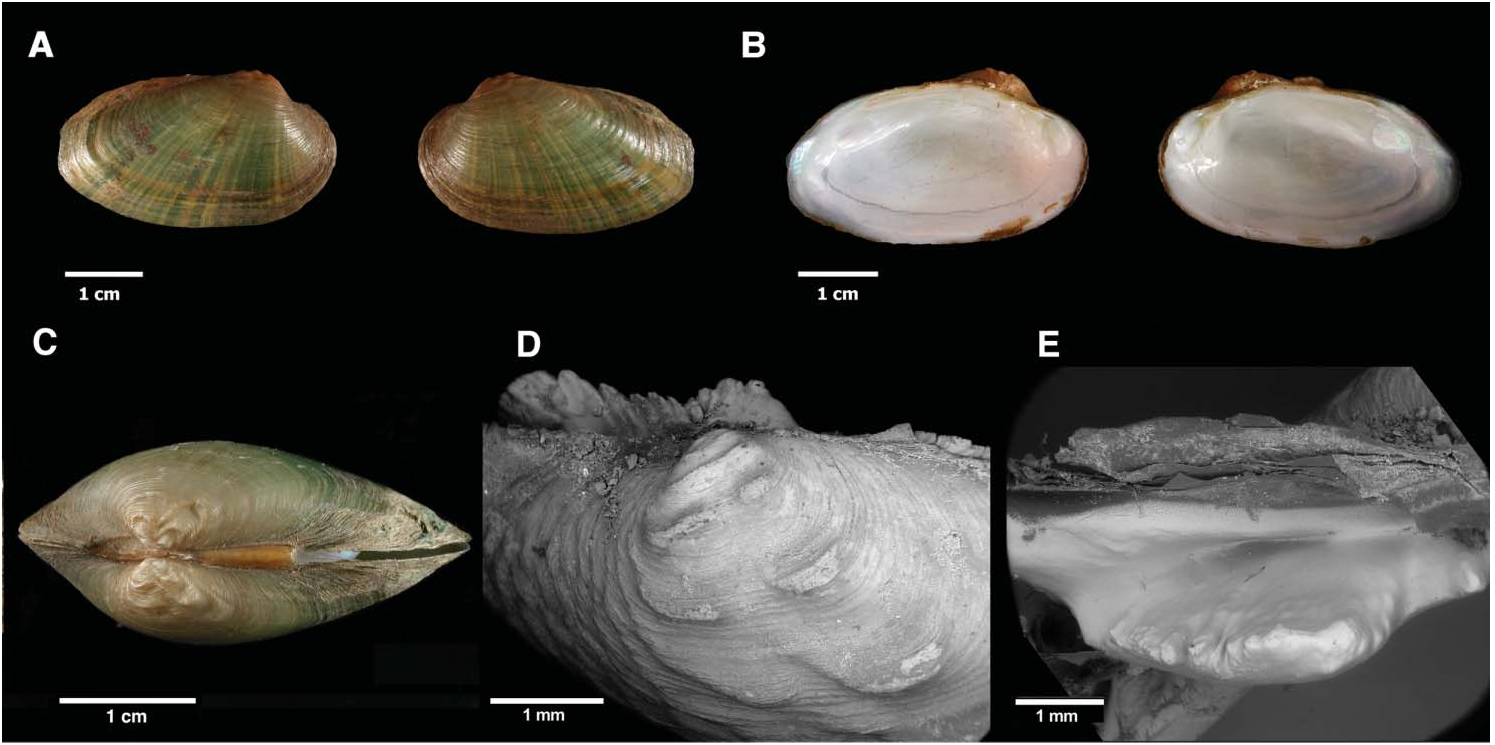
Shell equivalve and inequilateral with a regular oval outline, usually elevated at the posterior end ( Figures 2A,B View Figure 2 and 3 View Figure 3 ). In Ruidera lagoons ( Spain) often very rostrated or triangular in shape ( Figure 3C View Figure 3 ). Colour very variable, ranging from bright yellow to darkish green or brown ( Figure 3 View Figure 3 ). Often with a series of conspicuous yellow, green or black radial lines on the posterior end of the shell ( Figures 2A View Figure 2 and 3D View Figure 3 ). Umbo sculpture consisting of two or three rows of strong wavy rugae, followed by a variable number of successively weaker and less wavy rows. Rugae “w”-shaped or double “v”-shaped and parallel to the longitudinal axis of the shell ( Figure 2C,D View Figure 2 ). However, this pattern shows some variation and sometimes the typical “w” or double “v” rugae are not evident in the adult, often as the result of erosion of the shell, producing a sculpture with a more tubercular appearance. External ligament and hinge plate typical of Unio : left valve with two crenulated pseudo-cardinal teeth ( Figure 2D View Figure 2 ) and two laminar lateral posterior teeth variable in length; lower tooth always more elevated. Right valve with a small pseudo-cardinal tooth which is short, slightly curved with the concavity towards the dorsal side of the shell, and with an irregular serrated edge ( Figure 2E View Figure 2 ) and a laminar lateral posterior tooth variable in length. No supra-cardinal thickening in the right valve as in the Iberian Unio cf. pictorum . Maximum measured shell length 57 mm and maximum wet weight (shell + soft tissue) 16 g. Shell measurement ratios (mean and standard deviations) were as follows (with number of specimens analysed in parentheses): maximum length: maximum height (n = 92) 1.85 ± 0.09; maximum length: maximum width (n = 92) 2.57 ± 0.18; maximum length: anterior length (n = 25) 4.31 ± 0.35; maximum height: height at umbo (n = 25) 1.01 ± 0.03.
Anatomy
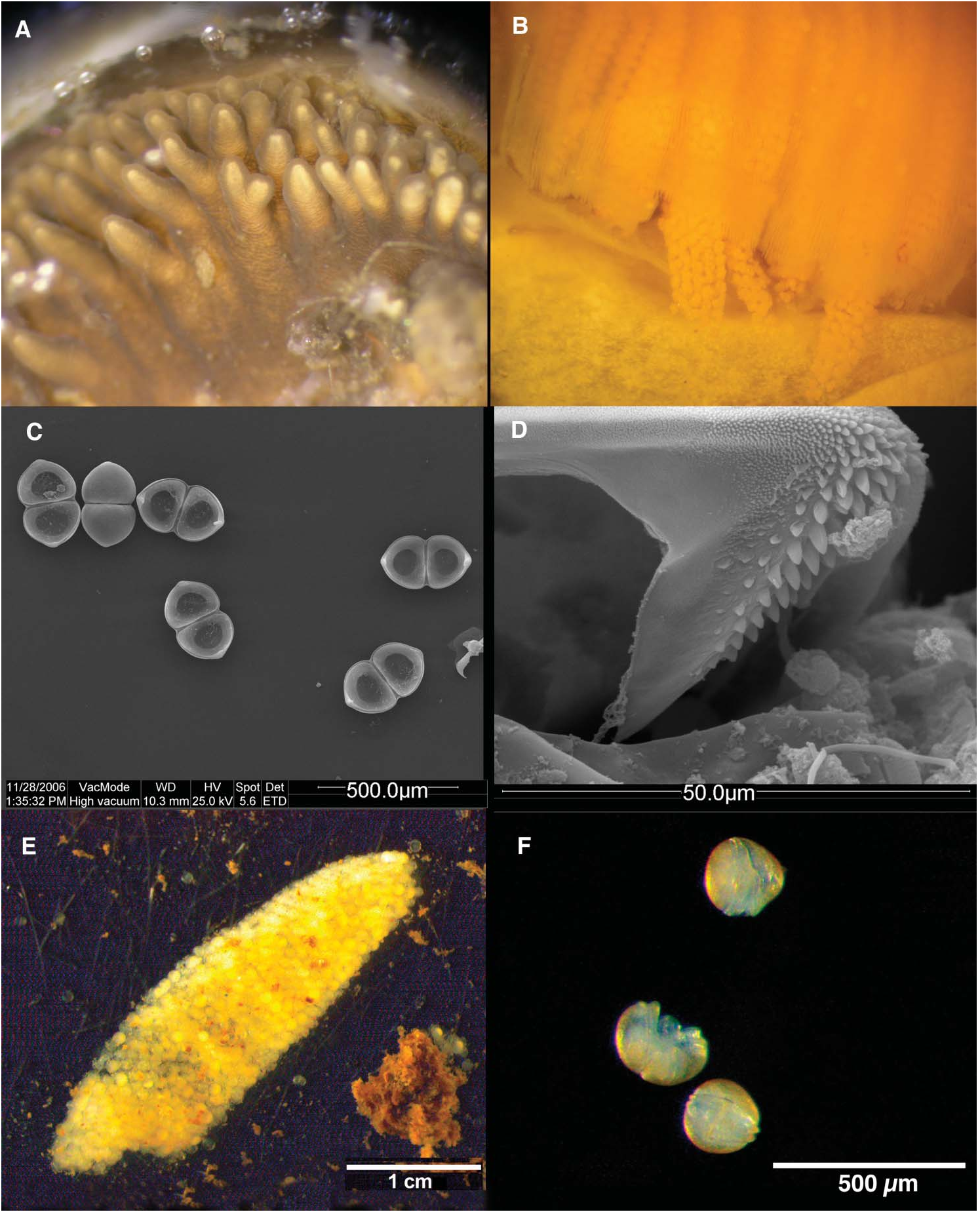
Soft parts under mantle lobes generally yellow-white in colour, although colour of the visceral mass (visible through the foot epithelium) variable depending on sex and season. Mantle-lobe margins fused in only one region (the bridge between the supraanal and the exhalant siphon), but three apertures visible in the posterior part, trimmed in brown-black and arranged from dorsal to ventral: the supra-anal, the exhalant and the inhalant siphons. Inhalant opening largest of the three openings and the only one surrounded by papillae; ventrally connected with the foot aperture (not a real siphon). One to three rows of papillae, sometimes bifurcated or with many branches ( Figure 4A View Figure 4 ). Space behind the inhalant and exhalant siphons separated by a diaphragm, not a mantle fusion. Diaphragm separates mantle cavity in the cloacal or suprabranchial cavity, and the branchial cavity below. Exhalant opening confined by two lateral muscular strips, formed by a row of not protruding papillae, with an external black trimming, and separated dorsally from the supra-anal aperture by a bridge built by the fusion of the inner mantle margins. Length of this bridge is shorter than that of the exhalant siphon, and shorter than the supra-anal aperture. Entire length of the outer lamella of the lateral demibranchs fused to the inner mantle wall. Inner gills only fused to the visceral sac anteriorly. After that point they run unattached and finally converge and join together behind the sac to form the diaphragm. Anteriorly, mouth bordered on either side by two white, rounded, undulating labial palps, which have a smooth external surface and a finely canaliculated inner side (facing each other).
Life cycle
Strictly dioecious species with no apparent sexual dimorphism. Zygotes incubated in the chambers formed by the water tubes until the glochidial stage ( Figure 4B View Figure 4 ). The entire internal cavity (homogeneity) of the external demibranchs acts as a marsupium (ectobranchy). Glochidia whitish, triangular in shape ( Figure 4C View Figure 4 ) and with a ventral styliform hook ( Figure 4D View Figure 4 ) that is armed with numerous spicules (teeth). Mean glochidial sizes of the population from São Pedro river (measured using scanning electron microscope): length 201.55 µm (SD = 4.83; n = 16), height 158 µm (SD = 3.54; n = 17), width 72.13 µm (SD = 4.07; n = 2). Glochidia release was only observed in captivity, with no evident stress factor associated. Mature glochidia released as a loose conglutinate with the shape of the brooding chambers ( Figure 4E View Figure 4 ) that sometimes includes eggs and embryos as well (indicating that the maturing of larvae is not always synchronous). No filament structure was observed in any conglutinate, which were very fragile and lost their shape very easily. Experiments with potential fish hosts resulted in juvenile drop-off ( Figure 4F View Figure 4 ) in aquaria containing Squalius alburnoides between 7 and 14 days after infestation at 26°C. No juveniles were found in aquaria containing other fish species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Unio tumidiformis Castro, 1885
| Reis, Joaquim & Araujo, Rafael 2009 |
Unio jourdheuili
| Azpeitia F 1933: 246 |
Unio conimbricus
| Kobelt W 1893: 99 |
Unio callipygus Drouët, 1893: 65
| Azpeitia F 1933: 243 |
| Drouet H 1893: 65 |
Unio baeticus
| Azpeitia F 1933: 254 |
| Kobelt W 1887: 55 |
Unio sadoicus
| Azpeitia F 1933: 249 |
| Locard A 1899: 261 |
| Castro JS 1885: 284 |
Unio macropygus
| Azpeitia F 1933: 251 |
| Locard A 1899: 262 |
| Castro JS 1885: 286 |
Unio eupygus
| Azpeitia F 1933: 252 |
| Locard A 1899: 262 |
| Castro JS 1885: 287 |
Unio batavus
| Morelet A 1845: 109 |