
Metaconchoecia alta
|
publication ID |
https://doi.org/ 10.5281/zenodo.156688 |
|
DOI |
https://doi.org/10.5281/zenodo.6276955 |
|
persistent identifier |
https://treatment.plazi.org/id/03E51C2A-FFD8-FFC3-FEE0-FD9383CDF990 |
|
treatment provided by |
Plazi |
|
scientific name |
Metaconchoecia alta |
| status |
|
Metaconchoecia alta View in CoL nov. sp.
( Figs.1525 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 View FIGURE 23 View FIGURE 25 )
Conchoecia skogsbergi: Chavtur, Shornikov, 1974: 286 (part)
Metaconchoecia skogsbergi: Chavtur, 1976: 105 View in CoL 106 (part); 1977a: 145146 (part); 1977b: 30 (part); 1977c: 20 (part).
Metaconchoecia View in CoL species nov. 3 Chavtur, 1991: 46 (part); 1992: 40, 49, table 2.
Etymology. The specific name “ alta ”, from the Latin adjective for high refers to height of the carapace.
Holotype. IBM 2800 – adult male, length 1.56 mm, appendages mounted on slide and valves in alcohol, in collection of the Museum of Institute of Marine Biology, Vladivostok, Russia (together, with paratypes).
Typelocality. RV Vityaz 39th Cruise, station 5617 (sample 126), 45º49’N; 153º33’E, depth layer 1000 500 m, August 5, 1966, using a Bogorov – Rass’s Net (S = 1,0 m²).
Paratypes. IBM 2801 – adult male, IBM 2802 – adult male, IBM 2803 – adult male, IBM 2806 – adult female, same station and sample as holotype; IBM 2804 – adult male, IBM 2805 – adult male, RV Vityaz station 6120 (sample 152) depth 750 500m, May 19, 1969; IBM 2807 – adult female, same station (sample 151) depth 1000 750 m. Paratypes IBM 28012803, 2806 – dissected, appendages and valves in alcohol: IBM 2804, 2805, 2807 – appendages on slide and valves in alcohol.
Additional material examined is listed in Appendix 2.
MALE
Carapace ( Fig. 15 View FIGURE 15 AF; Table 6 View TABLE 6 ). The carapace length of adult males specimens ranges from 1.371.65 mm (mean 1.53mm). The maximum height of the carapace is relatively high, being 4555% of the length, and occurs to the posterior of the midline, so that the posterior half of the shell is slightly higher than the anterior half. The greatest carapace width (4550% CL) is at, or just anterior to, the midpoint. The shoulder vault is weakly developed and rounded. The dorsal margin is straight, and joins the posterior end at a rounded angle with distinct apex. The posterior margin is evenly arched. The ventral margin is almost straight or gently curved and curves smoothly to join the posterior end. The rostrum is minute and has a pointed tip. The rostral incisure is quite shallow. In ventral aspect, the sides of the carapace are only gently curved. The right asymmetric gland opens close to the posterodorsal corner, and the left gland has opens somewhat behind the level of the incisure. The dorsomedial glands are weakly developed. There is no surface ornamentation.
Frontal organ ( Figs. 15 View FIGURE 15 G, 16 C). The shaft extends beyond the end of the second segment of the first antenna. The capitulum is expanded and spinose; the spines on the ventral surface are better developed. The ventral margin of the proximal half of the capitulum is strongly convex, whereas the distal part is either slightly tapered or parallel sided.
A and B – lateral view of left valve of shell, C – lateral view of right valve, DF ventral of shell, G – distal part of frontal organ.
A and B – upper lip, C – frontal organ and 1st antenna, D distal part of 1st antenna, E medial part of eseta of 1st antenna.
AC – 2nd antenna, D and E – endopodite of right 2nd antenna, F – distal part endopodite of left 2nd antenna, G – a and bsetae on endopodite of 2nd antenna.
A – endopodite of left 2nd antenna, B and C – anteroventral part of endopodite of left 2 nd antenna, D and E – anteroventral part of 1st endopodite segment of 2nd antenna, F – mandibule.
A – toothrows and masticatory pad of coxa of mandible, B – endopodite of maxilla, C – 5th limb, D – distal part of copulatory appendage.
A – basal endite of mandible, B – distal part of endopodite of maxilla, C and D endites of maxilla.
First antenna ( Fig. 16 View FIGURE 16 CE). The segmentation is fairly distinct. The eseta extends back parallel to the limb (except proximally where it loops down and is rather swollen) and barely reaches its proximal boundary. The bseta is slightly longer than the dseta. The cseta is thick, and shorter than the total combined lengths of segments 3 to 5. The most eseta is about 1.7 the length of the limb. The eseta armature comprises 1315 pairs of spines, which lie at an acute angle to the seta. The first segment has some dark pigment spots.
Second antenna ( Figs. 17 View FIGURE 17 AG; 18 AE). In some, but not all specimens examined the protopodite contains some dark pigment spots.The exopodite is about 60% the length of the protopodite. The first endopodite segment bears three triangular ridges, and the distoventral surface of the segment is covered with fine hairs. The processus mamillaris is bulbous with a beaklike extension pointing slightly forwards. The bseta is about twice the length of the aseta and 1.5 the length of the 2nd endopodite segment. The c and dsetae are relatively long. The eseta is large, stout and clawlike. The hook appendage on the right limb is strongly developed with a long curved distal section and a number of subterminal ridges; the corresponding appendage on the left limb is smaller, with a relatively short straight distal section rounded tip, and no subterminal ridges. The sensory h, j and isetae are about 40% the length of the gseta. The gseta is about 80% (or somewhat shorter) the length of the protopodite, about 1.3 the length of the exopodite and 4 times as long as the first endopodite segment. The f and gsetae are somewhat flattened distally.
Mandible ( Figs. 18 View FIGURE 18 F; 19 A; 20 A). The masticatory pad has four narrow rounded flaps (covered with exceedingly fine, rather short and exceedingly dense spines), 3 stout teeth, numerous filaments (with bifurcated tip) and hairs. The cutting edge of the basale carries two bare tube teeth; the posterior one is pointed and the anterior one rounded; these are followed by six teeth, the most posterior of which is bare whereas the others are serrated. The anterior inner tooth is triangular and serrated. The epipodite is triangular and developed. The exopodite is represented by relatively stout and plumose seta, which is somewhat longer than the length of the first endopodite segment. The dorsal seta on the first endopodite segment is plumose and rather stout being somewhat shorter than the segment itself. The most seta from the three dorsal setae on the second segment is rather stout and somewhat longer than the main clawlike seta of this limb.
Maxilla ( Figs. 19 View FIGURE 19 B; 20 BD). The most anterior seta on the endite of the precoxale is relatively long and distally bears thick secondary setae. Its main clawlike seta is bare. There are three posterior tube setae, one of which is armed with long spines, the other two are bare. On the posterior process of the coxal endite the most anterior tooth is bare. The basale has no seta. The first endopodite segment is somewhat narrow. The proportion between its width and the length is about 2 to 3. At about the middle of its anterior side are three shortly hirsute setae, and on the posterior side are four similar setae. The distomedial seta extends just beyond the end of the limb. On the base of this segment are about 10 short spines. The terminal segment is comparatively short and wide, and the anterior claw is about twice the length of the posterior claw.
A – 6th limb, B – terminal setae of 6th limb, C dorsal part of epipodite of 6th limb, D – copulatory appendage, E – copulatory appendage and furca.
A – apper lip, B – frontal organ and 1st antenna, C – 2nd antenna, D – endopodite of 2nd antenna, E – a and bsetae of endopodite of 2nd antenna, F and G – anteroventral part of 1st endopodite segment of 2nd antenna.
Metaconchoecia alta nov. sp. (Female: IBM 2801)
A – mandible, B and C – toothrows and masticatory pad of coxa of mandible, D – 7th limb.
Fifth limb ( Fig. 19 View FIGURE 19 C). The distal, middle and proximal groups of setae of the epipodial plate consist of four, four and four plumose setae respectively and the plate is also armed with few spines. The first endite of the protopodite bears one long plumose seta and another short shortly hirsute seta. The second endite carries a single long plumose seta and two short finely hirsute setae. On the endopodite there are eight setae, two of which are clawlike, twolong and plumose and remainder are relatively short and finely hirsute. The first exopodite segment bears seven ventral and one dorsal shorthaired setae and one lateral plumose seta. The second segment is with two ventral and one dorsal shorthaired setae. The end segment has two fairly long clawlike setae and a shorter ventral seta.
Sixth limb ( Fig. 21 View FIGURE 21 AC). The epipodial plate has five, five and six (and one additional short) long and plumose setae respectively in the distal, middle and proximal groups and is covered with some short spines. The longer seta on the endopodite is plumose. On the first exopodite segment there are three relatively short ventral, one plumose long lateral setae and distal row of hairs. The second segment has a minute ventral seta, and the third segment has a minute ventral seta and a longer dorsal seta lying parallel to the limb. The lengths of the terminal setae are subequal with the exopodite.
Seventh limb. The terminal segment bears two setae, one about 2.5 times the length of the other.
Caudal furca ( Fig. 21 View FIGURE 21 B). There are eight pairs of clawsetae and an unpaired seta dorsal to the shortest pair of clawseta. The inner surface is covered with fine hairs.
Copulatory appendage ( Figs. 19 View FIGURE 19 D; 21 D, E). The limb is relatively narrow and long ( Table 6 View TABLE 6 ). Its end is slightly truncated. The anterior side is straight (or muscle straight) near the end. There are five muscle bands.
Upper lip ( Fig. 16 View FIGURE 16 A, B). The posterior ventral edge is interrupted by a deep Vshaped notch. On each side of the notch there are some long flaccid spinelike processes, which decrease in strength medially.
FEMALE
Carapace ( Fig. 22 View FIGURE 22 AE). The length of adult specimens range from 1.401.67mm (mean 1.53mm). The maximum height of the shell is 5157% of the length, and the posterior sector is slightly larger than the anterior.
Frontal organ ( Figs.22 View FIGURE 22 FI; 23 B). The shaft extends well beyond the end of the first antenna and is about twice the combined length of the 1st and 2nd segments. The capitulum is separated from the shaft by undistinct suture. It is somewhat expanded proximally and tapers to a narrowly rounded end. Its dorsal surface is straight or slightly concave near the middle; the ventral surface is slightly concave or straight distally. The end is usually downturned. The surface of the capitulum is partially covered with minute spines.
First antenna ( Fig. 23 View FIGURE 23 B). The limb segmentation is fairly distinct. The first segment is bare, but on the second segment there is very small, bare dorsal seta. The a, b, c, dsetae have thick bases and are somewhat more than half the length of the eseta and are longer than the limb itself. The eseta is about twice as long as the limb and bears anterior and posterior spines in the distal half. It tapers to a pointed tip that is not flattened.
Second antenna ( Fig. 23 View FIGURE 23 CG). In some specimens the protopodite contains some dark pigment spots. The exopodite is short and about 65 % the length of the protopodite. The distoventral corner of the first endpodite segment bears three (rarely two) triangular ridges that are covered with fine hairs. The aseta is about one third the length of the bseta. The c (or? d) seta is absent. The f, g, h, i and j setae are different somewhat in length, but are more than half the length of the protopodite, about 2.5 as long as of the first endopodite segment, and about 80% (or somewhat more) the length of the exopodite, and have parallel sides and pointed or rounded tips. The processus mamillaris on the endopodite is similar in shape and size as in the male.
Mandible (Fig. 24 AC). There is no sexual dimorphism of this limb. The epipodite is wide, rounded and developed. The dorsal seta of the first endopodite segment is about 50% of the length of this segment.
Maxilla. Similar to the male’s.
Fifth limb ( Fig. 25 View FIGURE 25 A, B). Similar to that of the male, apart from having nine setae on the endopodite.
Sixth limb ( Fig. 25 View FIGURE 25 C). The limb is similar to that of the male, but differs in following respects: the first exopodite segment has five ventral and one dorsal plumose setae. The setae on the second and third segments are relatively long. The main terminal seta is subequal to the epipodial plate in length.
Seventh limb (Fig. 24 D), Caudal furca and Upper lip ( Fig. 10 View FIGURE 10 A) show little sexual dimorphism.
Remarks. This species is closely related to M. skogsbergi ( Iles, 1953) , but table 3 lists some of the characters that can be used to distinguish between the two species.
Distribution. This is a deep mesopelagic to bathypelagic species that occurs widely in the temperate waters of the North Pacific, the Sea of Okhotsk and the Bering Sea, but is absent from the Sea of Japan. Some specimens have been found in a single sample collected from the region of the Japan Trench (37ºN;143ºE). M. alta is collected between 37º and 61ºN at the depth range 200(300) 3000 m in the NorthWestern Pacific and between 51º and 60ºN from 300 to 4000 m in the NorthEastern Pacific ( Fig. 14 View FIGURE 14 ).
Its vertical distribution in the NorthPacific has been studied as an example of a planktonic species in the region of the KurileKamtchatka Trench (Table 7.). There it occurs over a the depth range of 3003000 m. Both its abundance and its biomass were greatest at 7501000 m, below that its abundance steadily declined to a particularly abrupt decline at 1500 m. The percentage contribution of M. alta to the numerous total halocyprid population is high; the maximum relative abundance (2532%) and biomass (1115%) were in the samples collected at 7501000 and 10001500m. Below 1500m their abundance and biomass decreased ten and thirtyfold respectively. Males and juveniles were most abundant at 7501000 m, and females were similarly abundant at 7501000 and 10001500 m layers ( Table 9 View TABLE 9 A).
In the Aleutian region M. alta mainly occurs at the depth range 3002500 m, although a single female was caught deeper (layer 38494002 m), but this may have been a contaminant. The maximum (mean) abundance and biomass were again in the 7501000 m, but were only half the abundances observed in the KurileKamchatka Trench, and again its abundance declined> 1000m. The contribution by M. alta to the overall halocyprid populations was similar to that observed in the North Western Pacific. Its contribution to the halocyprid population rose to a numerical maximum of 2127% and 1314% in terms of biomass at depths of 500750 and 7501000 m (Table 8). Once again below 1500m its percentage contribution to halocyprid numerical abundances dropped 5fold and 20fold in terms of biomass. Males were most abundant at 750 – 1000 m, females mainly occured at 500 – 1500 m, and juveniles at 500 – 1000 m ( Table 9 View TABLE 9 B).
TABLE 7. Abundance and biomass1 of halocypridids in area of the KurileKamtchatka Trench (RV Vityaz 39th Cruise, 1966; BogorovRass's Net, S=1,0 m2)․
Depth Num Total Halocyprididae Metaconchoecia alta sp. nov. ranges ber of
(m) catchs Abundance Biomass Abundance Biomass
(spec./10 3m 3) (mg/10 3m 3) (spec./10 3m 3) (mg/10 3m 3) 1 1 biomass is calculated on nomogrammes ( Chislenko, 1968) and equivalent to wet weight 2 ratio of mean abundance (or biomass) between M. alta nov. sp. and total Halocyprididae TABLE 8. Abundance and biomass1 of halocypridids in area of the Aleutian Trench (RV Vityaz 45th Cruise, 1969; Juday's Net Oceanic Model, S=1,0 m2)․
1 biomass is calculated on nomogrammes ( Chislenko, 1968) and equivalent to wet weight 2 ratio of mean abundance (or biomass) between M. alta nov.sp. and total Halocypridida males, females and juveniles in areas of the KurileKamtchatka (A) and Aleutian (B) Trenchs (RV Vityaz 39th
and 45th Cruises)
Depth ranges Male Female Juvenile Depth ranges (m) Male Female Juvenile (m)
0100 0100
100200 100200
200300 200300
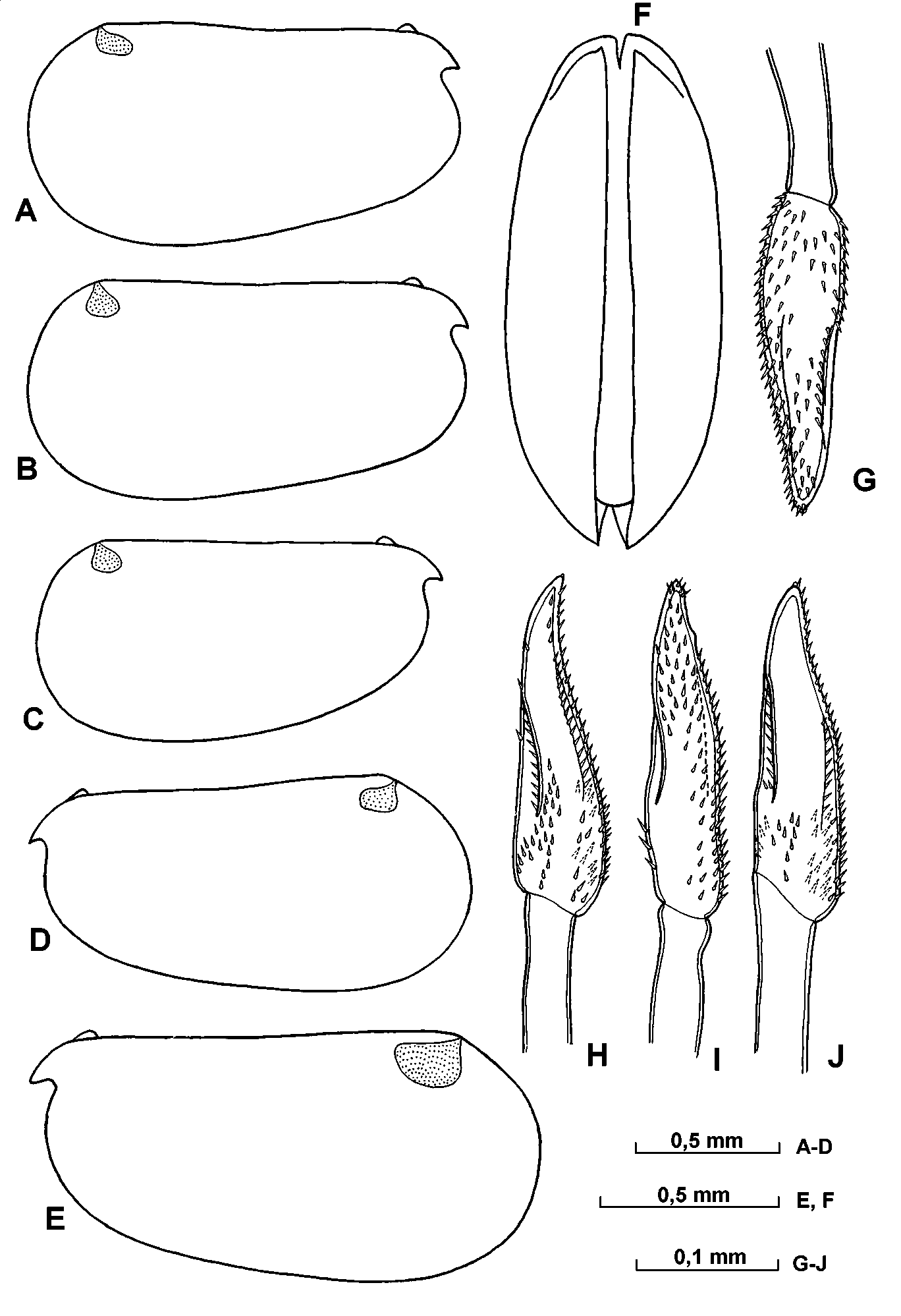
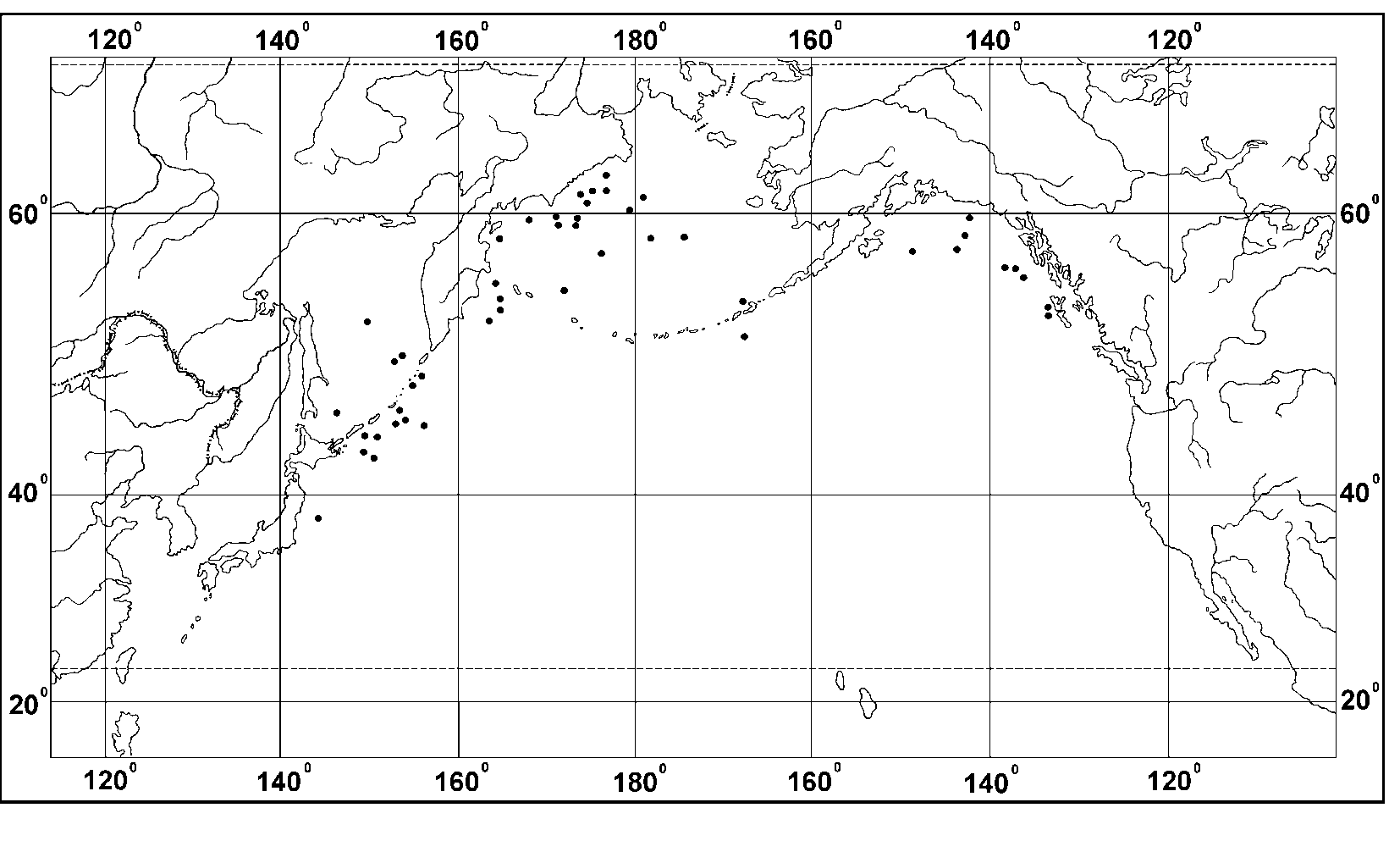
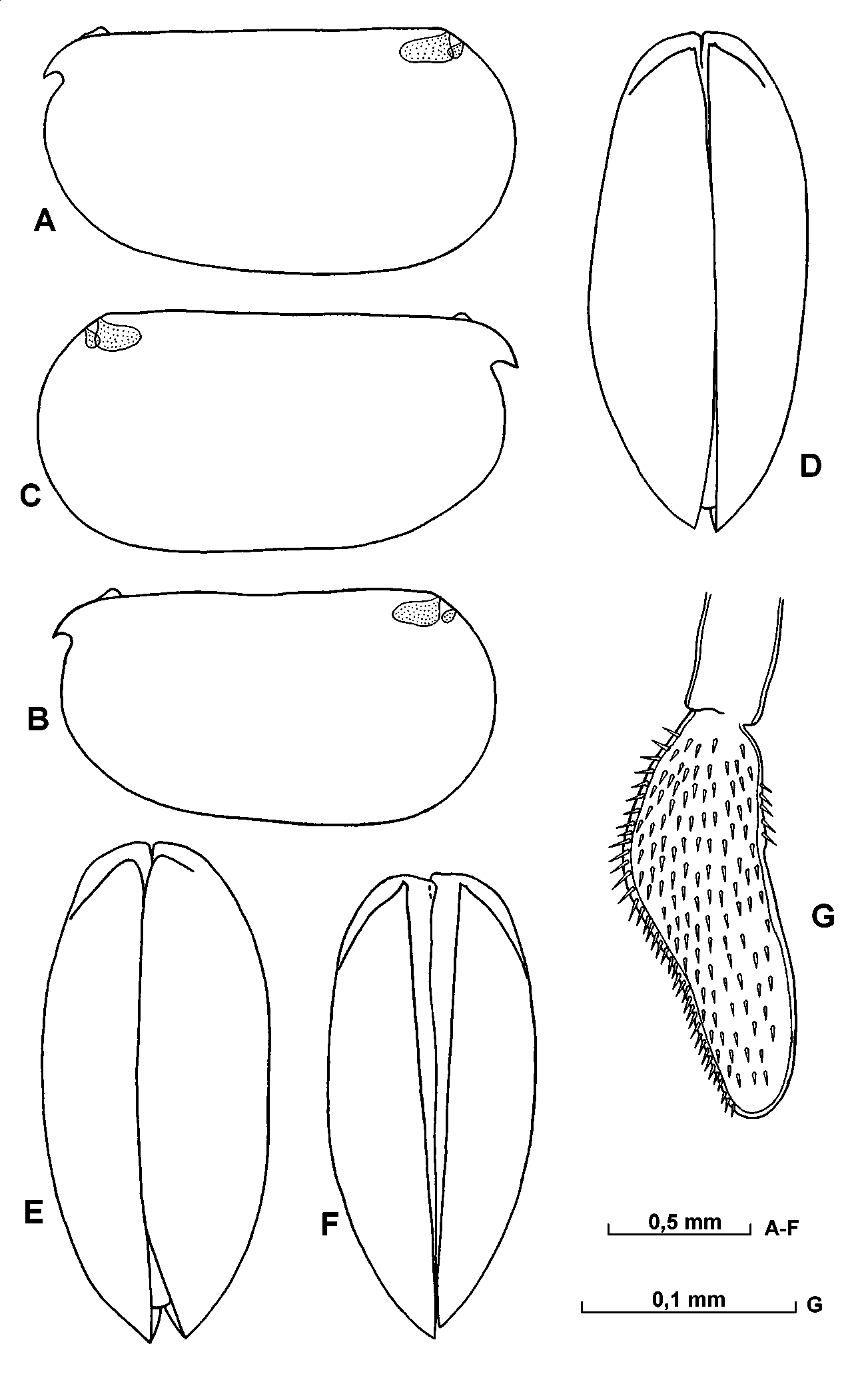
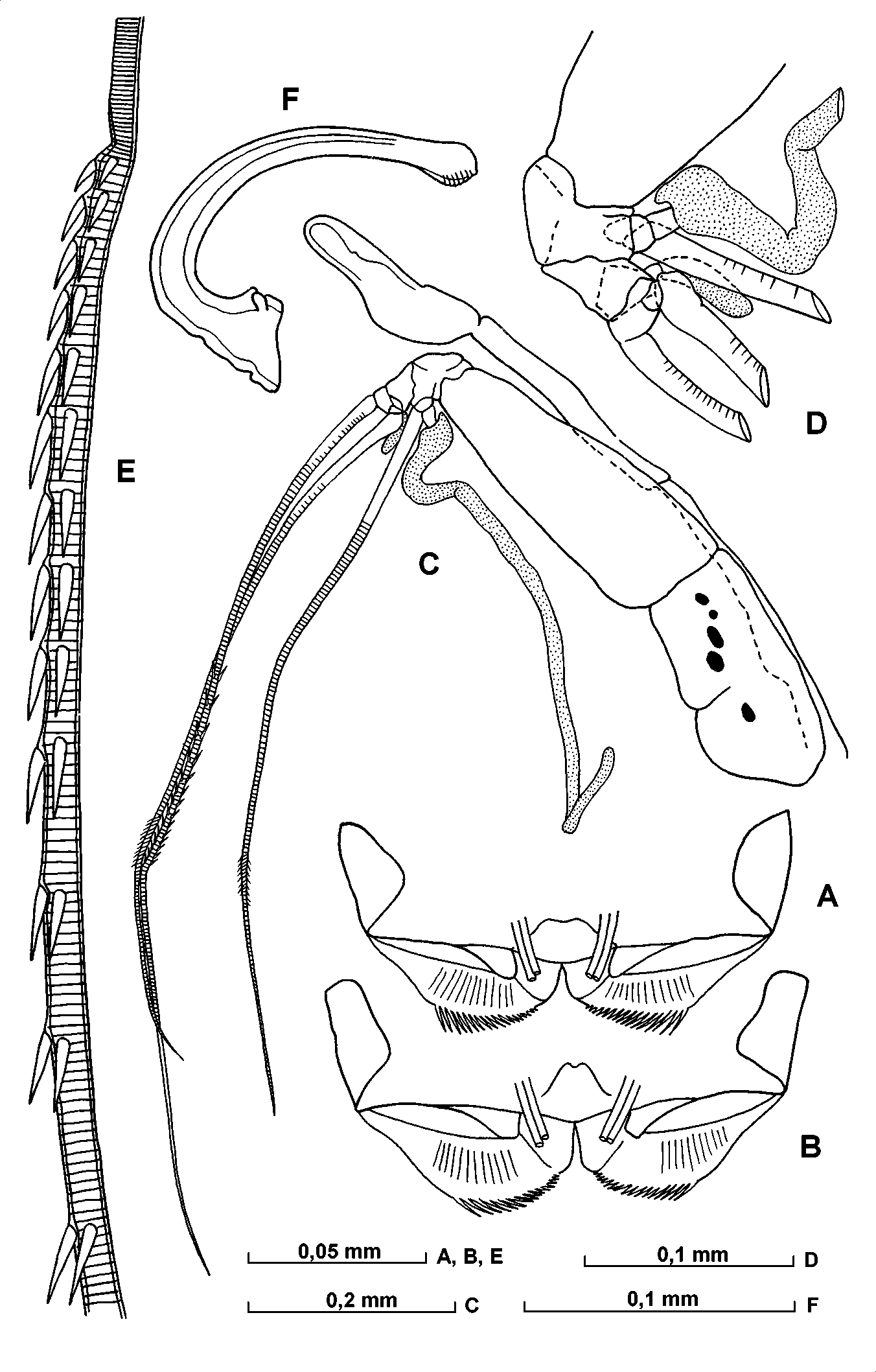
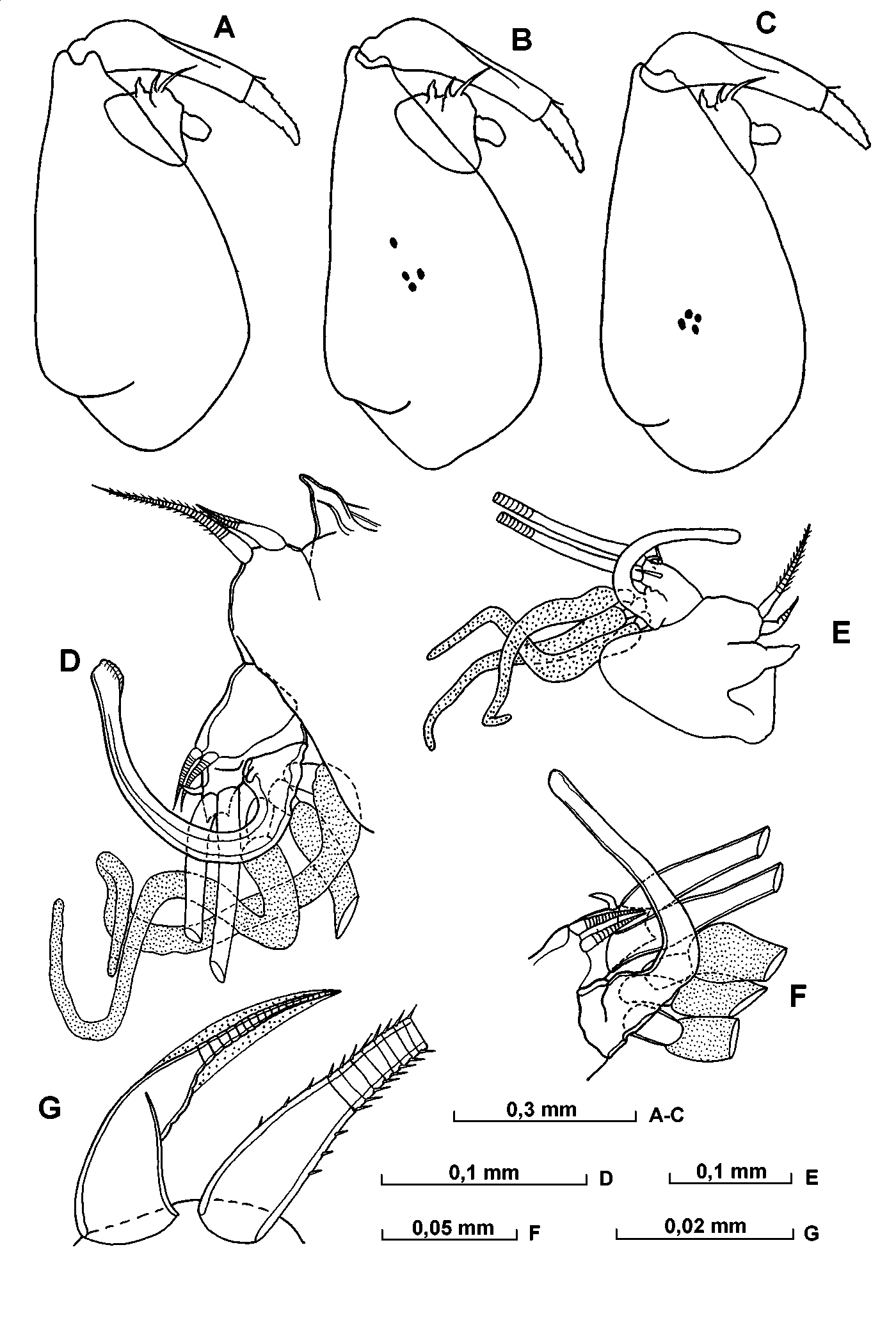
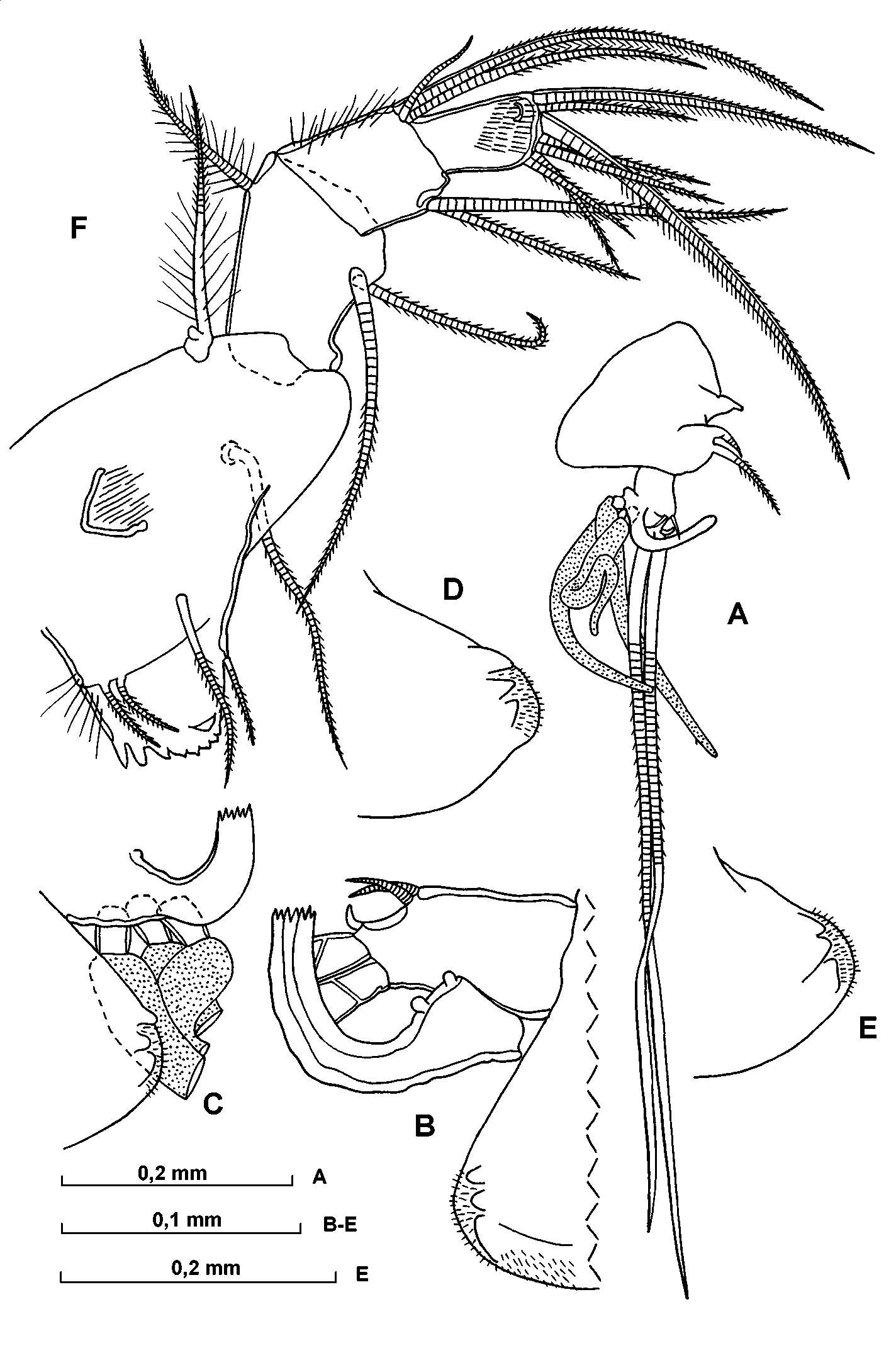
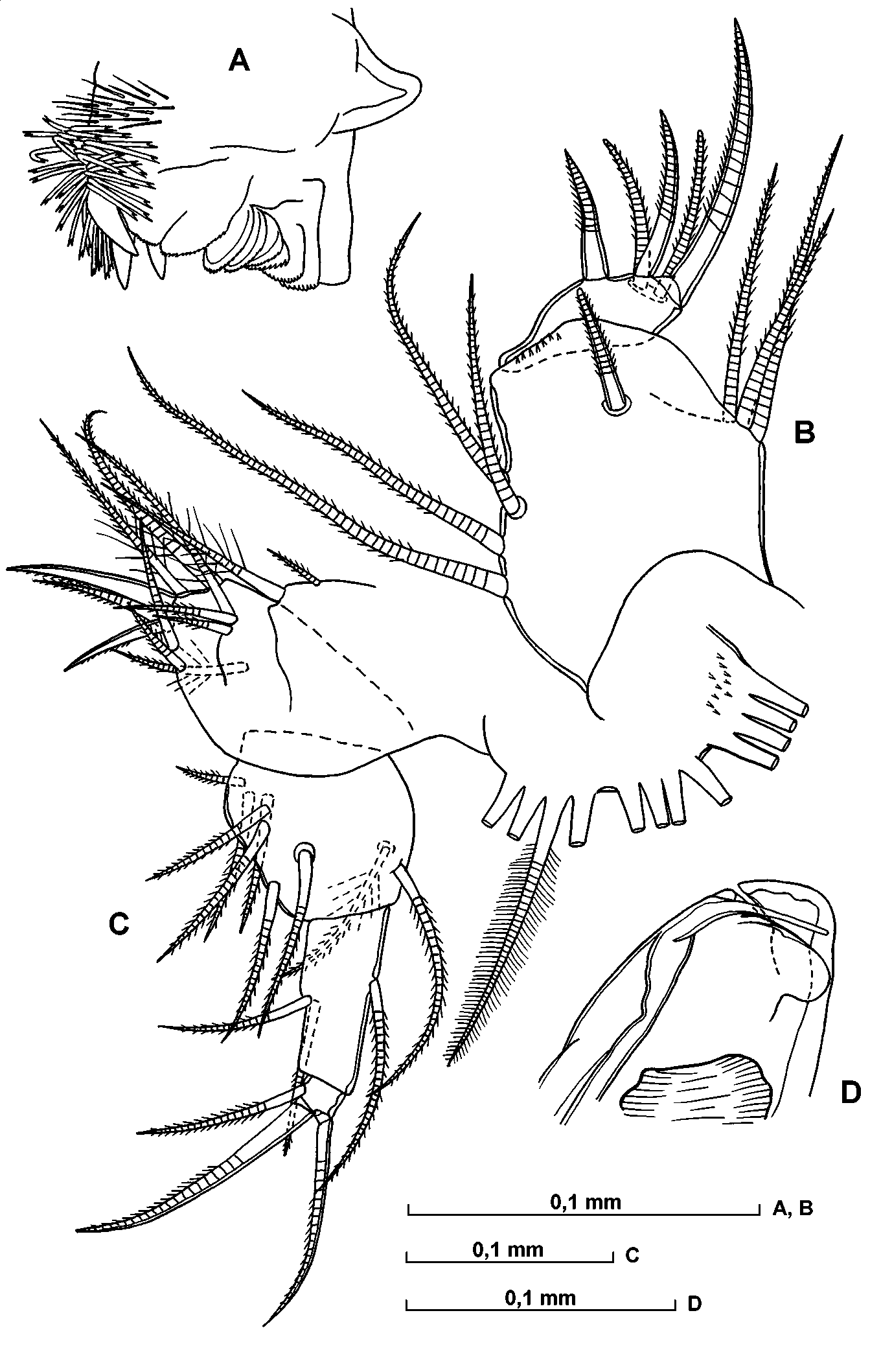
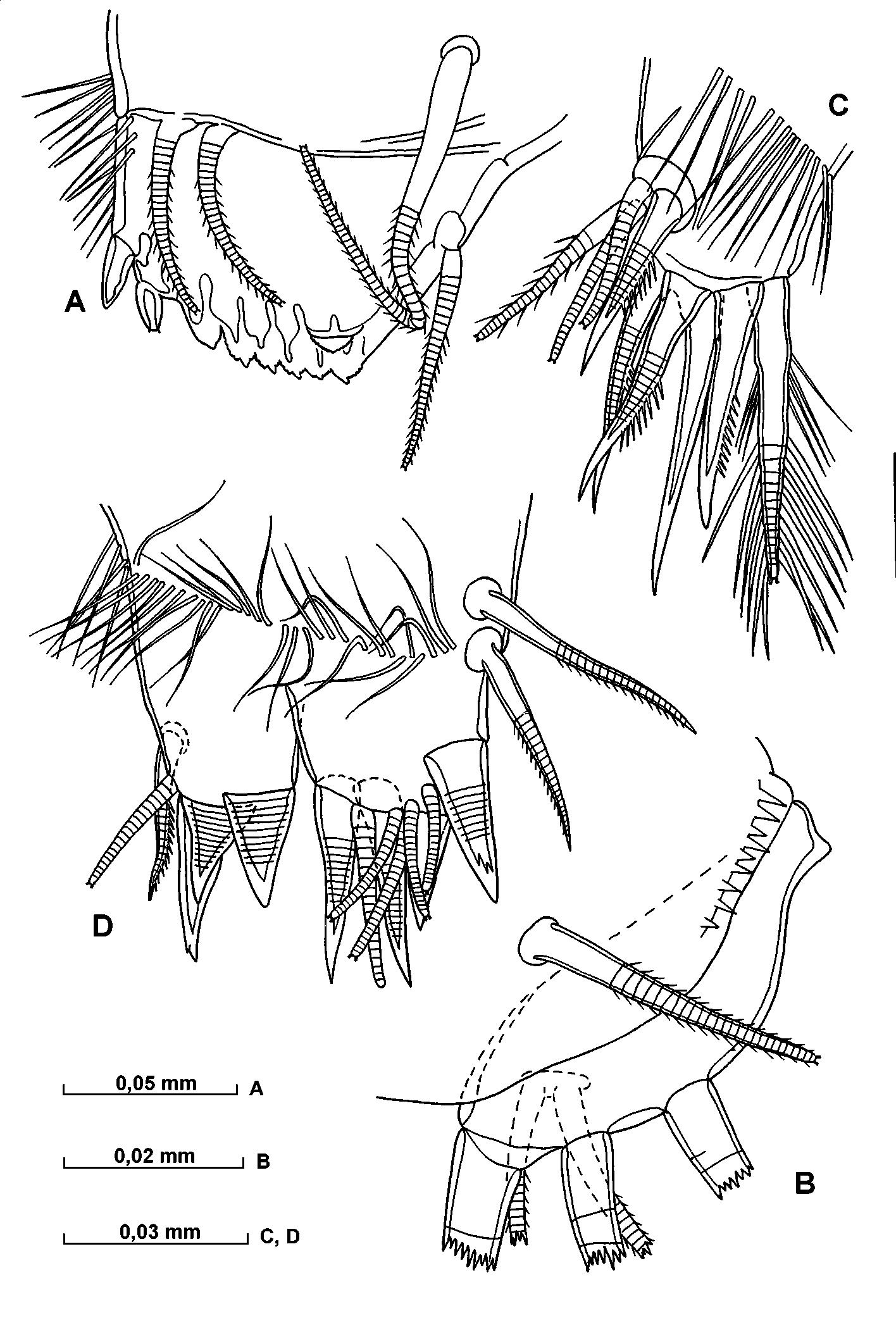
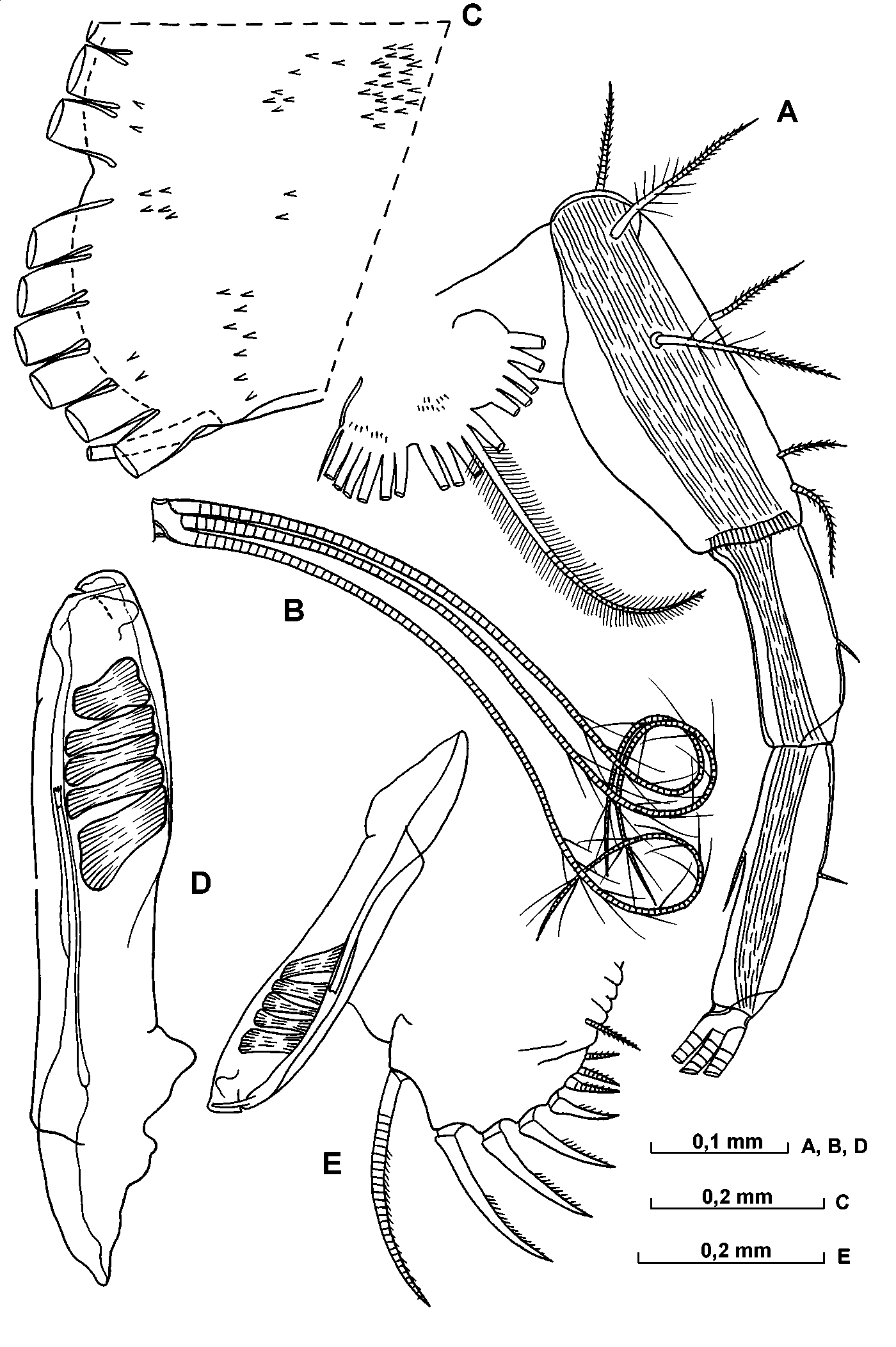
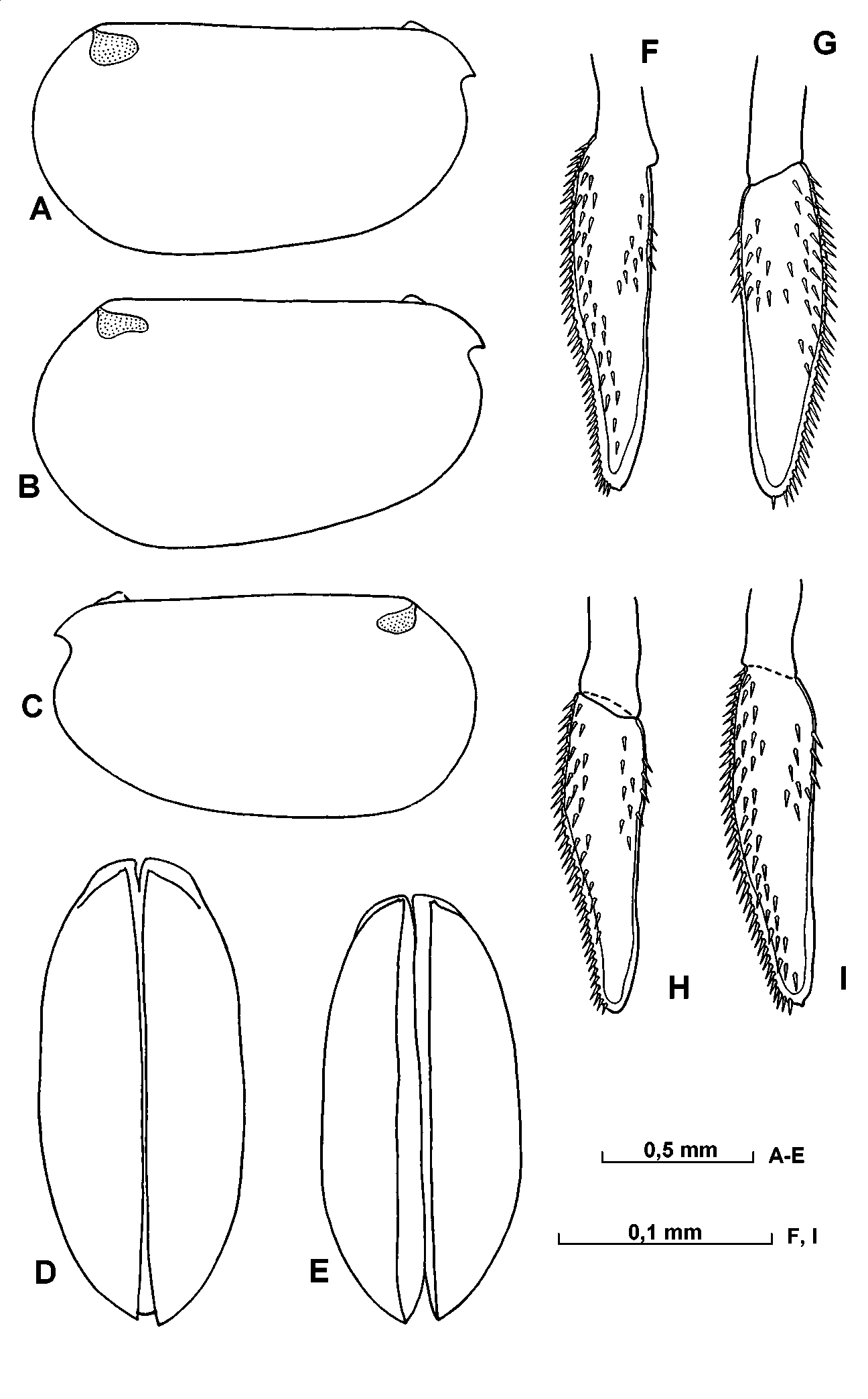
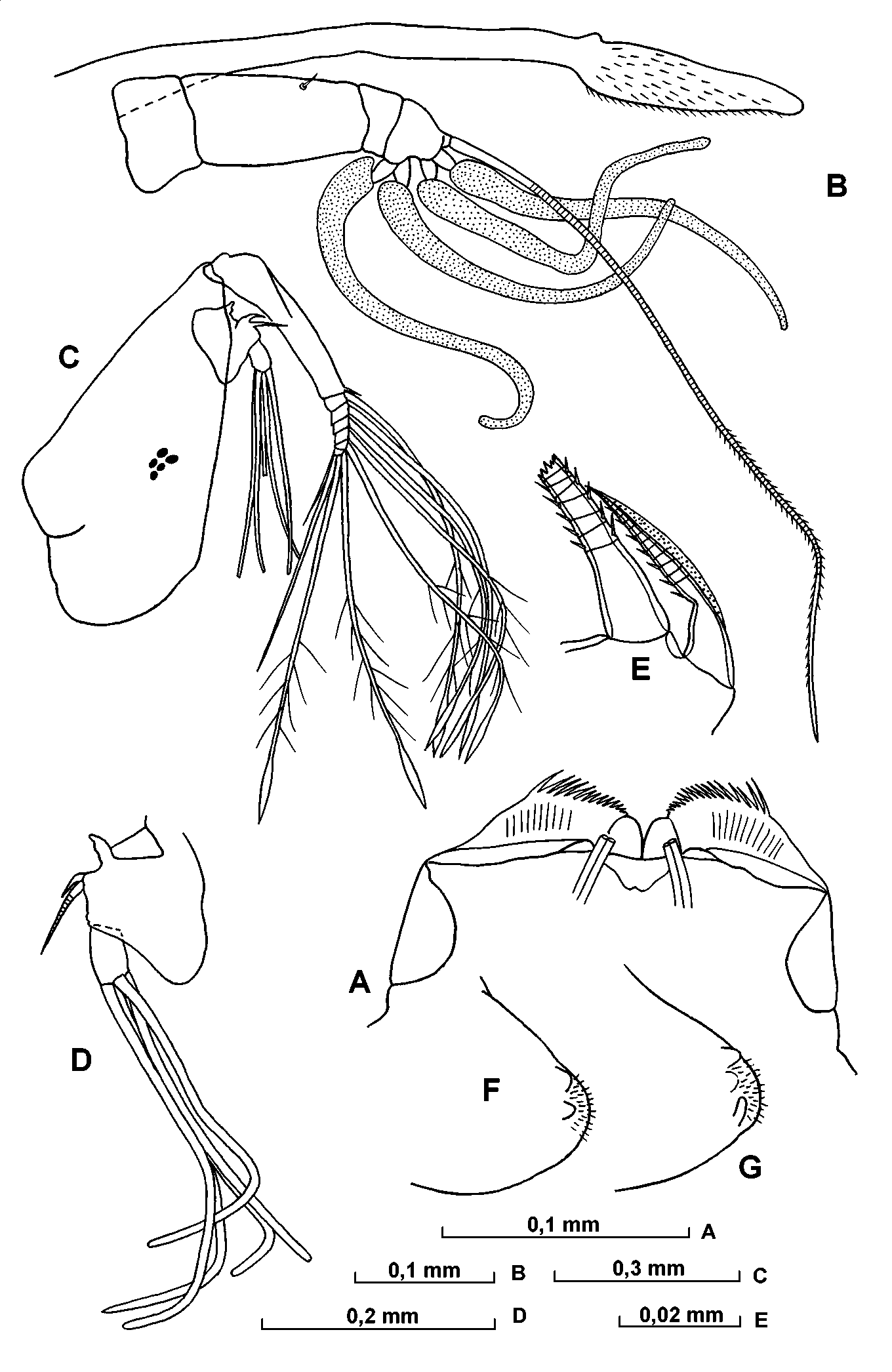
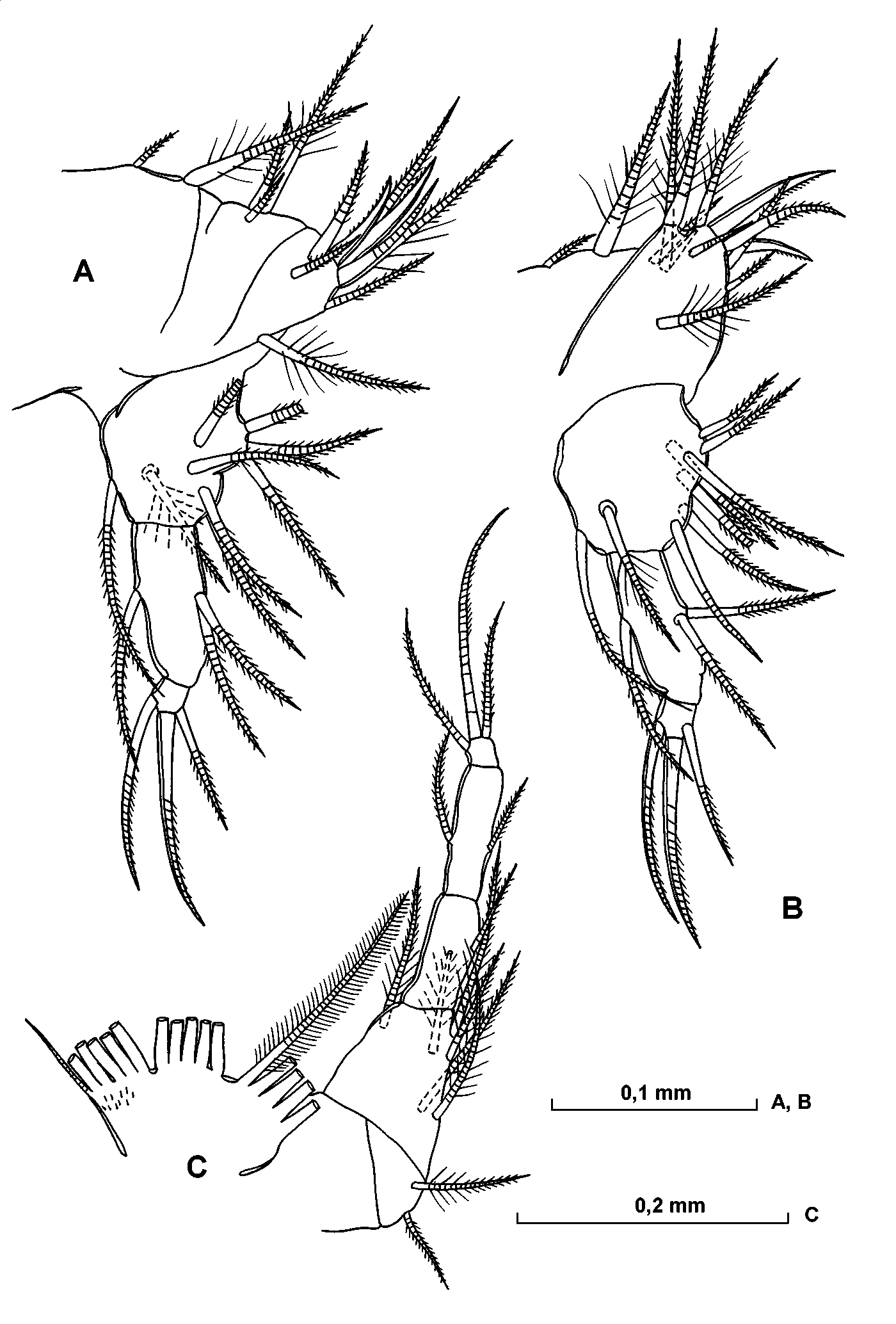
TABLE 6. Meristic characteristics of Metaconchoecia alta nov. sp.
| Characteristic | Range | Mean and number specimens |
|---|---|---|
| Length (mm): male | 1,371,65 | 1,53 (n=20) |
| female | 1,401,67 | 1,53 (n=22) |
| Height (%): male | 45,4555,33 | 51,42 (n=20) |
| female | 51,6156,67 | 54,90 (n=22) |
| Width (%): male | 45,1650,00 | 47,62 (n=20) |
| female | 42,7248,67 | 45,88 (n=22) |
| LAG (%): both sexes | 11,6113,33 | 12,54 (n=18) |
| Penis (% length of carapace) | 30,9735,33 | 32,93 (n=14) |
TABLE 9. Vertical profiles of densities (mean) of Metaconchoecia alta nov. sp. (numbers per 1000 m 3)
| Depth Num ranges ber of (m) catchs 050 8 | Total Halocyprididae Abundance Biomass (spec./10 3m 3) (mg/10 3m 3) Mean Range Mean Range 13.88 0 111 23.14 0 185 | Metaconchoecia alta sp. nov. Abundance Biomass (spec./10 3m 3) (mg/10 3m 3) Mean Range %2 Mean Range %2 (mean) (mean) |
|---|---|---|
| 50100 14 | 159.01 0 760 178.56 0 1557 | |
| 100200 26 | 210.74 01290 114.98 114.98 | |
| 200300 8 | 820.62 50 3029 567.01 83 1937 | |
| 300500 7 | 888.51 158 1824 840.26 329 1992 | 5.14 0 36 0.57 2.31 0 16 0.27 |
| 500750 3 | 180.9 75 276 128.06 63 161 | 49 22 87 27.08 17.98 Aug30 14.04 |
| 7501000 4 | 380.54 185 459 239.59 159 442 | 81.23 64 106 21.37 32.04 25 42 13.37 |
| 10001500 6 | 103.38 62 173 68.56 56 99 | 17.53 4 28 16.98 8.14 Feb13 11.87 |
| 15002000 5 | 43.21 20 90 105.65 11 110 | 1.49 0 4 3.44 0.7 0 2 0.66 |
| 20002500 4 | 29.52 13 46 144.46 94 254 | 0.9 0 4 3.05 0.36 0 1 0.25 |
| 25003000 3 | 57.5 19 89 74.18 21 116 | |
| 30003500 1 | 9.64 10 10.4 10 | |
| 35004000 2 | 64.63 16 113 49.06 Oct88 | 3.14 0 6 4.85 1.76 0 4 3.59 |
| 40005000 0 | | |
| 50006000 0 | | |
| 60007000 1 | 6 6 4 4 | |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Metaconchoecia alta
| Chavtur, Vladimir Grygorjevich 2003 |
Metaconchoecia
| Chavtur 1991: 46 |
Metaconchoecia skogsbergi:
| Chavtur 1976: 105 |