
Phelsuma astriata
|
publication ID |
https://doi.org/10.5281/zenodo.194693 |
|
DOI |
https://doi.org/10.5281/zenodo.5615793 |
|
persistent identifier |
https://treatment.plazi.org/id/03E487B6-FFA8-FFD0-D5CB-95C1FDACFDE4 |
|
treatment provided by |
Plazi |
|
scientific name |
Phelsuma astriata |
| status |
|
Phelsuma astriata group
(contains P. astriata and P. sundbergi )
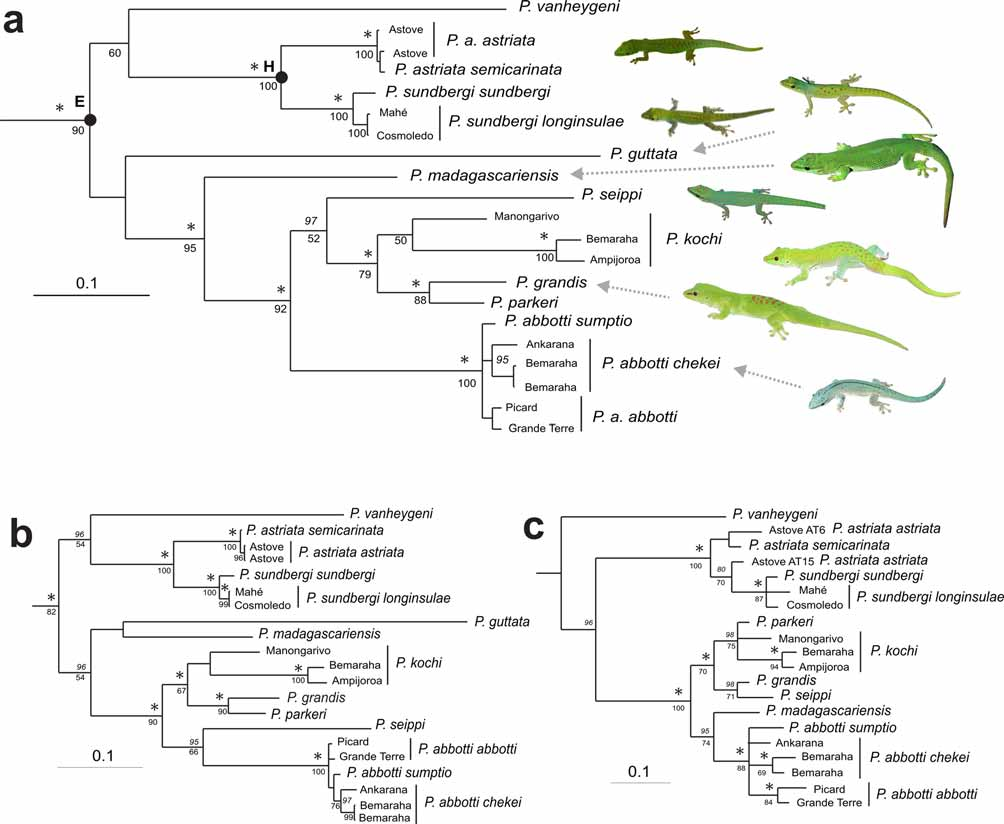
These two species form a small endemic radiation on the Seychelles islands and various subspecies have been described for both. Variation within this Seychellois group was studied by Cheke (1982), Gardner (1984, 1986, 1987), and more recently, by Radtkey (1996). The available molecular data, nuclear and mitochondrial, is unambiguous in supporting monophyly of this group ( Fig. 5 View FIGURE 5 : clade H), which furthermore was recovered in single-gene analyses of all genes except C- mos, where differentiation relative to other species in clade E (see Fig. 5 View FIGURE 5 ) is very low. The mitochondrial and combined analyses indicate that, possibly, Phelsuma vanheygeni is the sister species of the Seychellois clade and may be in the future included in the P. astriata group (see section on this species below).
Species in the Phelsuma astriata group are all green coloured with a weak dorsal pattern and no or only weak dorsal or lateral lines, keeled dorsalia and lateralia, size-reduced vertebral scales, and are non-gluers, but otherwise appear to be quite variable in morphological characters ( Tables 1–2 View TABLE 1 ).
Relationships within the P. madagascariensis group
This group is in a well-supported clade ( Fig. 5 View FIGURE 5 ; clade E) with the Seychellean P. astriata group, and with P. vanheygeni . The basal topology of clade E is uncertain; the position of both P. vanheygeni and P. guttata is unresolved, mainly because of conflicts between the mitochondrial and nuclear datasets. In the mitochondrial, as well as in the combined data, P. vanheygeni is sister taxon to the P. astriata group and P. guttata is placed in the P. madagascariensis group. The nuclear data, however, place P. vanheygeni basal in clade E, and P. guttata completely outside of it, albeit both placements with less than 50% PP/BS support. Because of its morphological affinities to the P. madagascariensis group we nevertheless include P. guttata in this group, but we emphasize that further study on its phylogenetic position is needed.
Within the P. madagascariensis group, the P. guttata species group as previously defined ( P. g u t t a t a, P. abbotti , P. masohoala , and P. seippi ; Glaw & Vences 1994; Glaw et al. 1999; Van Heygen 2004) does not form a monophyletic group; instead, P. guttata is placed basal in the group, and P. s e i p p i and P. abbotti also do not appear to be sister species. Two members of this group originated by (long) distance colonization in a northwestwards direction: P. abbotti abbotti (Aldabra) and P. parkeri (Pemba) . Both have their closest relatives in the northwestern/northern Malagasy coast, the most likely area in fact from which colonizations to occur from, considering surface sea-currents in the region ( Ali & Huber 2010).
One of the few nominal Phelsuma species for which no DNA sequences are available until now is P. masohoala . As discussed by Meier & Böhme (1996) it is possible that P. masohoala is related to P. abbotti and thus belongs to the P. madagascariensis group where we provisionally include it.
Relationships among P. madagascariensis , P. grandis , and P. k o c h i
In our molecular analysis, Phelsuma grandis , P. kochi , and P. madagascariensis which were long considered subspecies of a single species ( P. madagascariensis ) did not form a clade, although the mitochondrial and nuclear data suggested different relationships among these taxa. This confirms the results of Raxworthy et al. (2007) who recently proposed to elevate them to species level, based on data from a molecular analysis and from environmental niche modelling. Neither Raxworthy et al. (2007) nor ourselves had sequences of Phelsuma madagascariensis boehmei Meier, 1982 , and the status of this taxon thus remains unclarified.
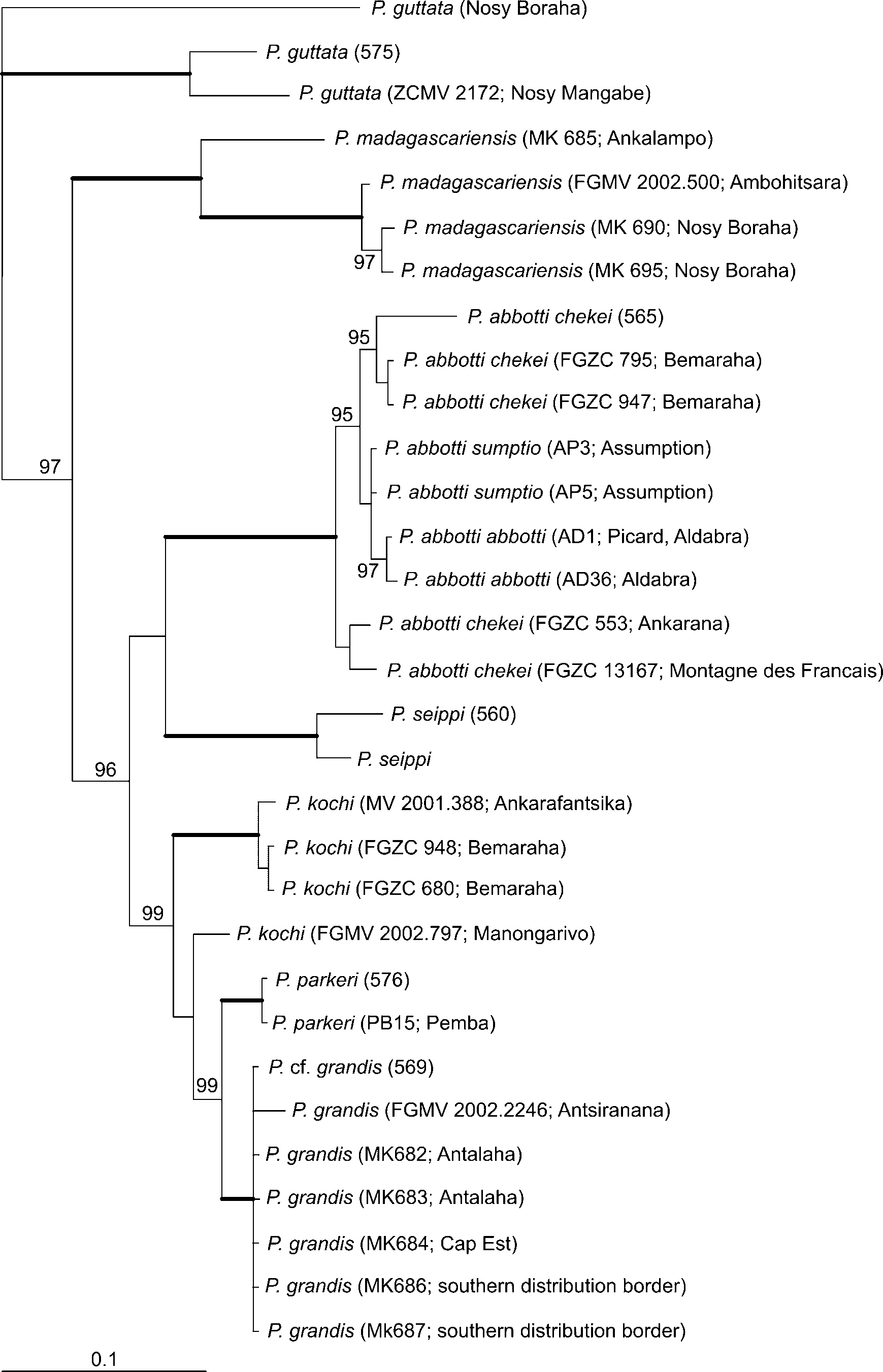
We detected deep genetic divergences among P. grandis , P. kochi , and P. madagascariensis , reaching up to 14% (uncorrected p-distances from cytochrome b). Within P. grandis and P. kochi distances of up to 1.1% and 9% were, respectively, observed for cytochrome b. These surprisingly high distances prompted us to undertake a wider survey of variability in the P. madagascariensis group, based on mitochondrial 16S rDNA sequences. The three taxa, P. grandis , P. kochi , and P. madagascariensis are large geckos that are thought to be allopatrically distributed mainly in coastal Madagascar: P. madagascariensis in the east, P. grandis in the north, and P. kochi in the west ( Meier & Böhme 1991), with contiguous distributions and contact zones between them. The new data ( Figs. 5 View FIGURE 5 and 6 View FIGURE 6 ) add various new perspectives to this picture:
First, we detected a second deep lineage that we here assign to P. kochi in a preliminary way: a specimen (identified as P. k o c h i) from Manongarivo in the Sambirano region, far northwestern Madagascar, differed strongly from specimens from the west (Tsingy de Bemaraha and Ankarafantsika). For 16S alone, it did not even form a monophyletic group with the remaining P. k o c h i ( Fig. 6 View FIGURE 6 ), and although it did in the combined analysis ( Fig. 5 View FIGURE 5 ), support was not significant (PP<50%; BS=50%).
Second, also samples of P. madagascariensis were deeply differentiated, with an eastern-southeastern clade (from Ambohitsara and Nosy Boraha) and a north-eastern clade (from Ankalampo), and with a differentiation between these of 7.5% uncorrected p-distance in the 16S gene ( Fig. 6 View FIGURE 6 ).
Differentiation within P. abbotti
A last aspect obvious from the multi-gene phylogeny ( Fig. 5 View FIGURE 5 ) and the 16S data ( Fig. 6 View FIGURE 6 ) is the low differentiation among the various subspecies of Phelsuma abbotti . In Madagascar, the subspecies P. a. chekei shows a slight differentiation between specimens from the north (Ankarana and Montagne des Francais) and west (Tsingy de Bemaraha). Phelsuma a. abbotti and P. a. s u m p t i o from Aldabra and Assumption islands show some differentiation but certainly have colonized these islands very recently, as expected.
Phenotypic characterization of the P. madagascariensis group
Species in the Phelsuma madagascariensis group are characterized by non-gluing egg laying behaviour, smooth ventral and subcaudal scales, keeled dorsal and lateral scales absence of nostril-rostral contact, and size-reduced vertebral scales, whereas other characters are variable among species in the group, possibly in part related to the size differences ( Tables 1–2 View TABLE 1 ). For example, the number of preanofemoral pores is higher in the larger species ( P. grandis , P. kochi , P. madagascariensis , P. parkeri ). Several characters such as nongluing behaviour, and absence of rostral-nostril contact are shared by the P. madagascariensis group and the P. astriata group, in accordance with their placement in a monophyletic group by the molecular data, but P. vanheygeni which also belongs into this clade is an egg-gluer (see below).
Definition and content of the group
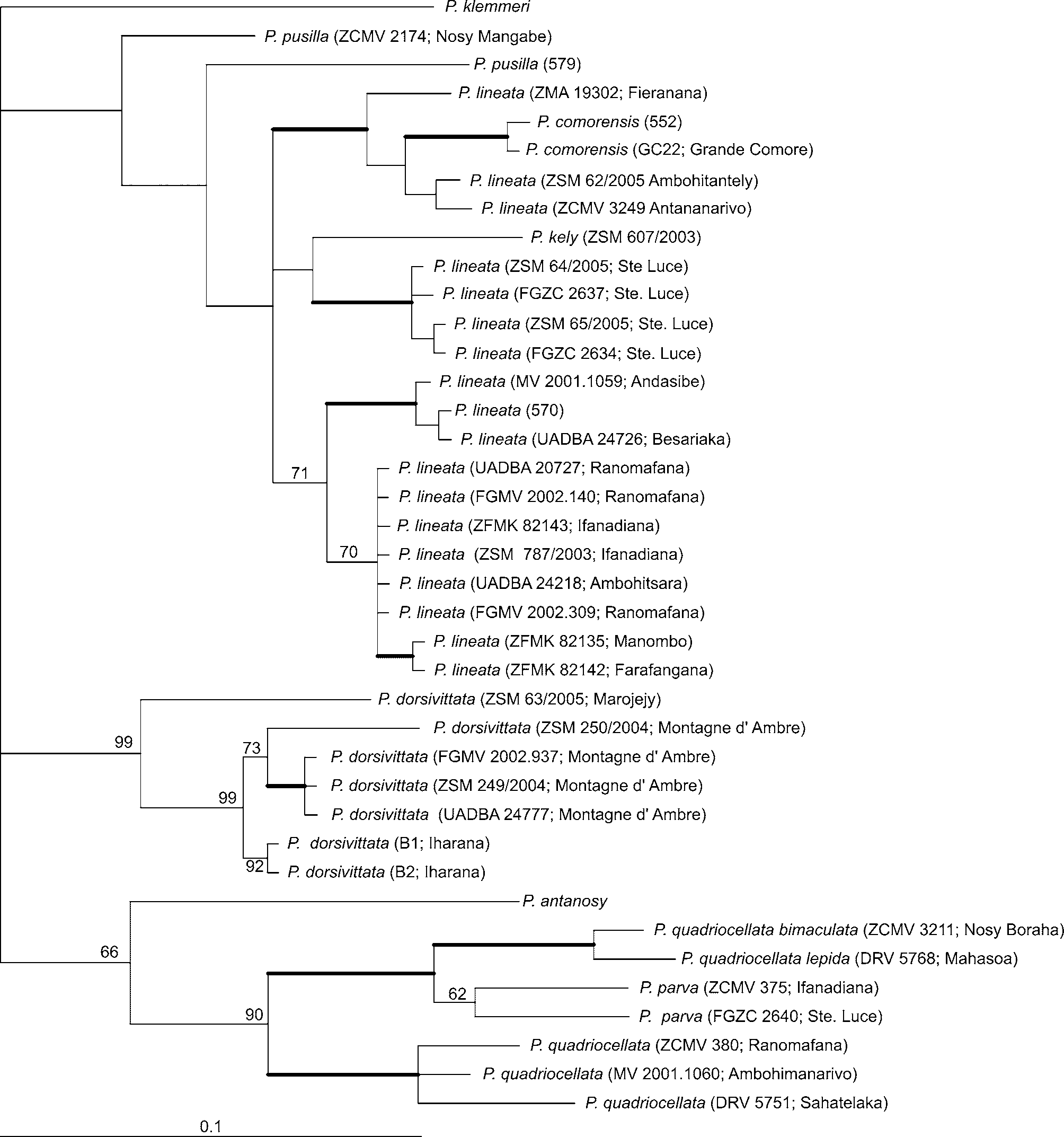
Molecular data strongly corroborate the previously suggested ( Glaw & Vences 1994) evolutionary relationships between P. lineata , P. quadriocellata and P. pusilla . Other species belonging to this well supported group are P. k e l y, the Comoran P. c o m o re n s i s, and P. antanosy ( Fig. 7 View FIGURE 7 a; clade J) Two subgroups can further be defined: (1) all P. quadriocellata subspecies (including P. quadriocellata bimaculata , sometimes regarded as a distinct species – Glaw & Vences, 1994) and P. antanosy and (2) P. lineata , P. pusilla , P. k e l y and P. c o m o re n s i s. Although nuclear genes reveal unresolved or paraphyletic relationships between these ( Figure 7 View FIGURE 7 c), for the faster evolving mtDNA markers these groups already reached reciprocal monophyly and are recovered with strong support ( Fig. 7 View FIGURE 7 b).
Phelsuma antanosy was previously thought to be more closely related to either the species from the Mascarenes ( Raxworthy & Nussbaum 1994), i.e., the P. cepediana group as defined here, or to the P. laticauda group ( Glaw & Vences 1994) but in the molecular analysis it is clearly nested within the P. lineata group, sister to P. quadriocellata .
Although no molecular data are yet available, we here assign the recently described ( Berghof & Trautmann 2009) Phelsuma hoeschi to the P. lineata group based on its dark lateral stripe and its similarities to P. pusilla and P. kely in scale characters, as also emphasized in the original description (see also Tables 1– 2 View TABLE 1 ).
Loveridge (1942) hypothesized about the dark spots of P. quadriocellata being derived from the dark lateral band of P. lineata . It is worth mentioning that P. l. bombetokensis , a geographically isolated taxon from dry forest in north-western Madagascar (still unstudied from a molecular perspective), has an intermediate pattern in this respect.
Variation within the P. lineata complex and recognition of P. dorsivittata at species level
Phelsuma lineata , being paraphyletic in respect to P. k e l y, P. c o m o re n s i s and P. p u s i l l a (if lineata is considered as including dorsivittata as subspecies), exhibits consistent deep differentiation (12.8% uncorrected p-distances for cytochrome b) between P. lineata dorsivittata from the far north of Madagascar (including a specimen of uncertain status from Marojejy in north-eastern Madagascar) and the remaining individuals. Genetic distances within remaining P. lineata individuals for cytochrome b reach 7.7% uncorrected p-distances. Also, p-distances reach as much as 8.8% within P. lineata dorsivittata . The expanded analysis based on 16S sequences only, but including many more specimens (those sequenced by Boumans et al. 2007, plus 16 others) largely confirms these conclusions ( Fig. 8 View FIGURE 8 ). Phelsuma kely and P. c o m o re n s i s are deeply nested within the P. lineata clade. A taxonomic consequence from this fact could be to accept a paraphyletic species P. lineata or, given the known colour differences between populations ( Krüger 1996c), its partitioning into various species. The following deeply differentiated clades can be distinguished: (1) P. lineata populations from the highlands (Antananarivo, Ambohitantely, Fierenana), which would correspond to P. lineata elanthana according to Krüger (1996c), (2) populations from the south east (Ste. Luce), (3) populations from the northern central east and southern central east (Andasibe, Besariaka, Ranomafana, Ifanadiana, Ambohitsara, Farafangana), and (4) as the most divergent lineage, the populations from the north, currently corresponding to P. l. dorsivittata . However, P. lineata sensu lato is almost continuously distributed over most of Madagascar's rainforests, with probable contact and possibly hybrid zones, especially between population groups from the highlands and the northern central east. Furthermore, the nuclear genes provide no evidence for separating lineages 2 versus 3 ( Fig. 7 View FIGURE 7 c; highland specimens of lineage 1 not included). The available evidence is therefore insufficient for conclusive statements on the taxonomic status of the lineages 1–3. On the contrary, lineage 4 from the northern localities Montagne d'Ambre, Iharana (= Vohemar) and Marojejy is consistently recovered by mitochondrial and nuclear markers ( Fig. 7 View FIGURE 7 b–c) and is grouped basal to the clade containing P. l i n e a t a lineages 1–3, P. pusilla , P. kely and P. c o m o re n s i s ( Fig. 7 View FIGURE 7 a). We see this as sufficient support for a status of the northern populations as separate species, Phelsuma dorsivittata .
Typical specimens of Phelsuma dorsivittata have several red spots on the back, which can often fuse to larger markings. Usually a red vertebral stripe is present on the anterior back. A rather indistinct dark lateral spot is present behind the forelimb, and can be also present in front of the hindlimbs, forming the anterior and posterior part of a dark lateral line (distinctly recognizable in the holotype). However, the anterior spot is not isolated and not bordered by blue colour in life. In Phelsuma dorsivittata 8–9 supralabialia (min-max) and 6–9 infralabialia (min-max) are present. Selected morphological characters of the male holotype (SMF 59373, Fig. 9 View FIGURE 9 ) of Phelsuma dorsivittata are as follows: SVL 55.3 mm; TL 124.4; 9/9 supralabials on the left and right side of the head; 8/7 infralabials; 1 internasal; nostril in contact with rostral and first supralabial; 28 preanofemoral pores; mental triangle-shaped; dorsals, ventrals, and subcaudals keeled. For a more detailed description of the holotype see Mertens (1964). The holotype, upon examination in 2009 and based on the original description, shows the colour pattern as described above (distinct anterior dark marking on the flanks, red dorsal markings forming a midventral line on the anterior part of the back. However, a specimen from Marojejy (the same as included in our molecular trees) as well as another specimen from the type locality Montagne d'Ambre (see Glaw & Vences 2007: p. 403 and their fig 1f) show a rather uniform dorsum with no or few red markings which do not form a longitudinal line. Whether this variation is related to the substantial molecular variation observed among these populations and among specimens from Montagne d'Ambre requires further study.
Variation within the P. quadriocellata complex and recognition of P. parva at species level
Within P. quadriocellata , uncorrected p-distances for cytochrome b reach 13.4% and three main clades seem to be distinguishable by the mtDNA markers ( Figs. 7–8 View FIGURE 7 View FIGURE 8 ), although nuclear markers do not provide resolution (and show evidence of ancestral polymorphism) at this level. One of these likely corresponds to typical P. quadriocellata and appears to be distributed over much of the mid-altitude localities in the northern central east and southern central east, from Sahatelaka (near Lake Alaotra) to Ranomafana. A second clade was found in the lowlands of the southern central east and south east (Ifanadiana and Ste. Luce). The specimens from these two sites are distinctly smaller and have a different pattern of red dorsal markings than those from nearby Ranomafana, and thus corresponded to the current subspecies P. quadriocellata parva . Specimens from lower elevations in the northern central east ( Nosy Boraha and Mahasoa) clustered together (uncorrected p-distance for the 16S gene 2.2%) ( Fig. 8 View FIGURE 8 ) and correspond morphologically to two subspecies: specimens from Mahasoa possibly to P. q. lepida and specimens from Nosy Boraha possibly to P. q. bimaculata .
Given that especially quadriocellata and parva occur in close geographic proximity (Ranomafana vs. Ifanadiana) but each of them is genetically and morphologically constant over wide areas of eastern Madagascar ( parva: Ste Luce in the southern central east to Ifanadiana in the southern central east; quadriocellata: Ranomafana in the southern central east to Sahatelaka in the northern central east; Fig. 8 View FIGURE 8 ), we hypothesize that these two taxa are distinct at the species level, and consider Phelsuma parva as full species. Average cytochrome b distances between P. q. quadriocellata and P. parva are high (11.9%), and average distances within P. p a r v a and P. q. quadriocellata are high as well, 7.3% and 5.6% respectively, for cytochrome b.
Phelsuma parva is one of the smallest species within the genus Phelsuma ; its total length does not reach more than 90 mm ( Meier 1983; Glaw & Vences 2007). The body has a slender shape with a bright green colouration. It has distinct red spots on the back and a dark spot behind the forelimb that is variable in size, more or less round and often surrounded by a blue ring. Phelsuma parva is characterised by 7–10 supralabialia (min-max), 7–8 infralabialia (min-max). Selected morphological characters of the male holotype (ZFMK 19315, Fig. 9 View FIGURE 9 ) of Phelsuma parva are as follows: SVL 36.2 mm; TL 80.6; 7/7 supralabials; 7/7 infralabials; 1 internasal; nostril in contact with rostral and first supralabial; 22 preanofemoral pores; mental triangle-shaped; dorsals, ventrals, and subcaudals keeled. For a more detailed description of the holotype see Meier (1983). All specimens collected by us and included in our molecular trees agree in size and colour closely with the holotype.
Specimens assigned to the newly defined species Phelsuma parva do not form a monophyletic group in the analysis based on nuclear DNA sequences ( Fig. 7 View FIGURE 7 c). However, the same would also be true for P. quadriocellata sensu lato, if parva was seen as subspecies of such an inclusive species. Moreover, the fact that rather well-defined species such as P. antanosy , P. comorensis , P. pusilla , and P. kely cluster among individuals of P. parva and P. l i n e a t a in the nuclear gene tree indicates that these placements are probably due to incomplete lineage sorting in these complexes of species.
The recognition of P. parva at species level also rises questions about the status of individuals here assigned to P. q. bimaculata and P. q. lepida which in the mitochondrial analysis form a clade with P. parva rather than P. q. quadriocellata ( Fig. 7 View FIGURE 7 ). These forms have distinct differences in colour patterns from each other, from P. q. quadriocellata , and from P. p a r v a. We have little doubts that they are to be assigned to one or possibly even two separate species. However, the nomenclatural situation is convoluted because (1) a clear definition of the taxon bimaculata based on a re-examination of type material is missing, and (2) our analysis does not include topotypical samples of lepida (which was described from near Andapa in the North-East), so that the identity of this taxon requires confirmation too. We therefore here refrain from taxonomic conclusions and postpone these to forthcoming studies.
Phenotypic characterization of the P. lineata group
The Phelsuma lineata group is characterized by a predominant green colour with usually a pattern of lateral/sublateral stripes, typically with keeled ventral and subcaudal scales (except P. lineata punctulata P. lineata elanthana and P. c o m o re n s i s), absence of rostral-nostril contact, a relatively low number of midbody scales (below 98, except for P. antanosy ) and absence of size-reduced vertebral scales ( Tables 1–2 View TABLE 1 ). The P. lineata group shares with the P. laticauda group (see below), which is its sister group ( Fig. 7 View FIGURE 7 ), the non-gluing behaviour and absence of nostril-rostral contact, but species in the P. laticauda group have smooth ventral and subcaudal scales, and several of them have size-reduced vertebral scales.
The P. lineata group as defined here is one of the genetically most diverse clades within Phelsuma , and will probably turn out to be the most species-rich clade after a thorough taxonomic study. We had no P. lineata punctulata or P. lineata bombetokensis available but considering their isolate distribution (in central North and North-West Madagascar, respectively) it is possible that these taxa will again turn out to be genetically differentiated from the remaining.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.