
Daspletosaurus torosus, Russell, 1970
|
publication ID |
https://doi.org/10.5281/zenodo.1040973 |
|
DOI |
https://doi.org/10.5281/zenodo.4704932 |
|
persistent identifier |
https://treatment.plazi.org/id/03E3C818-5444-5328-FEB0-C9933752FD12 |
|
treatment provided by |
Jeremy |
|
scientific name |
Daspletosaurus torosus |
| status |
sp. nov. |
Daspletosaurus torosus new species *
Distribution
Oldman Formation, near Steveville, Alberta, except where noted.
Type
NMC 8506 skull and skeleton, lacking hind limbs (quarry 88 of Sternberg 1905).
Paratype
AMNH 5438 View Materials sacrum and adjacent thoracic and caudal vertebrae, pelvis, right femur, left tibia and second metatarsal ( Little Sandhill Creek basin).
Referred Specimens
NMC 350 left hind Hmb (sec. 31, tp. 20, rge. II, W. 4th mer.).
AMNH 5346 View Materials maxilla (Little Sandhill Creek basin).
UA 11 right femur and fourth metatarsal.
NMC 11594 incomplete, weathered skull and lower jaws (Oldman Formation, near Manyberries, Alberta: near centre NW.;!: sec. 36, tp.l, rge. 6, W. 4th mer., about 30 feet below prairie rim or 3,285 feet above sea level).
BMNH R4863 View Materials premaxilla, maxilla, dentary.
Diagnosis
Premaxilla does not contact nasal below external naris. Maxillary teeth large, not greatly reduced posteriorly. Anteriormost antorbital fenestra within maxilla minute, hardly visible in lateral aspect, first antorbital fenestra nearly as high as long. Nasals slightly constricted between lacrimals, invaded posteriorly on either side by long slender tongue of frontal. Frontal broadly exposed on skull roof, lacks deep vertical cleft above orbit. Prefrontal broadly exposed dorsally between nasal, lacrimal, and frontal. Lacrimal ' horn' triangular in lateral aspect, with apex centred above antorbital ramus of bone; exceeds postorbital ' horn' in development. Anteroexternal edge of antorbital ramus of lacrimal is not continued across lateral surface of jugal. Ventral process of extopterygoid inflated, with large ventrally opening sinus; jugular process of bone also inflated. Angular terminates posteriorly beneath centre of surangular foramen.
Ventral opening of main basisphenoid sinus situated on midline of skull, behind transverse wall linking basipterygoid processes. Optic fenestra not bridged medially by presphenoid. Exits for cranial nerves III and IV contained in single pit on anterior surface of laterosphenoid. Exit for cranial nerve Vi situated on lateral surface of laterosphenoid.
Neural spine of fourth cervical vertebra much larger than that of fifth. Neural spine of fifth cervical distally pointed, that of sixth cervical smaller and sharply pointed. Presacral vertebrae tend to be shorter and higher than in Albertosaurus ; basal caudal vertebrae diminish less rapidly in size posteriorly than in latter species.
In adults length of humerus estimated to equal 38 per cent of that of femur, forelimb powerfully developed relative to that of Albertosaurus . Anterodistal condyle of metacarpal I reduced, causing first digit to diverge distally from second. First phalanx of digit II robust in adults. Dorsal edge of ungual of digit I passes through arc of about 90 degrees.
Anterior blade of ilium covers diapophysis of twelfth thoracic vertebra. In adults circumference of femur equal to 38 to 41 per cent of length of femur. In comparison with Albertosaurus specimens of similar femur length, presacral vertebral column is much shorter, thoracic ribs are slightly longer, forelimb is longer, ilium and sacrum are longer, and metatarsus is slightly shorter.
Comments
The type specimen of D. torosus was collected by C. M. Sternberg late in the 1921 field season. As is usual in large theropod specimens, the head and anterior part of the vertebral column were pulled dorsoposteriorly toward the pelvis before the animal was buried. Although the hind limbs and many of the more distal caudal vertebrae were lost and although the remainder of the skeleton was partly scattered, the specimen is generally very complete and very well preserved. Sternberg's field notes indicate that at the time the specimen was collected he thought it belonged to an undescribed species of Gorgosaurus or to a new genus. Photographs of the skeleton in situ have been published by Sternberg (1945: 191; 1946: pi. 15; 1966: fig. 9), and of the skull by Sternberg (1945: 191; 1946: pi. 6; 1966: fig. 17).
Daspletosaurus attained approximately the same length (28-30 feet) as its contemporary Albertosaurus . However, the body, which measured 1,350 mm (53 inches) through the widest point of the thoracic region in NMC 8506, and the base of the tail are much heavier than in Albertosaurus , and the hips and hind limbs are also more powerfully developed. The forelimb, although small, is relatively larger than in any other known tyrannosaur. Interestingly, with the exception of NMC 350, which is subadult, only fully grown specimens of Daspletosaurus have so far been collected from the Oldman Formation, while remains of one-half to three-quarters grown albertosaurs are relatively not uncommon in these sediments. The simultaneous existence of these two large carnosaurs indicates the presence in the fauna of two écologie niches to accommodate them. As the endocranial cavities of both forms are quite small, it might be expected that differences in their gross morphology would rather accurately express their behavioural differences. Perhaps the more powerfully constructed daspletosaurs preyed on ceratopsians, and the more fleet albertosaurs were better suited to subsist on hadrosaurs. Albertosaurs and hadrosaurs are indeed correspondingly more abundant in the Oldman fauna than are daspletosaurs and ceratopsians.
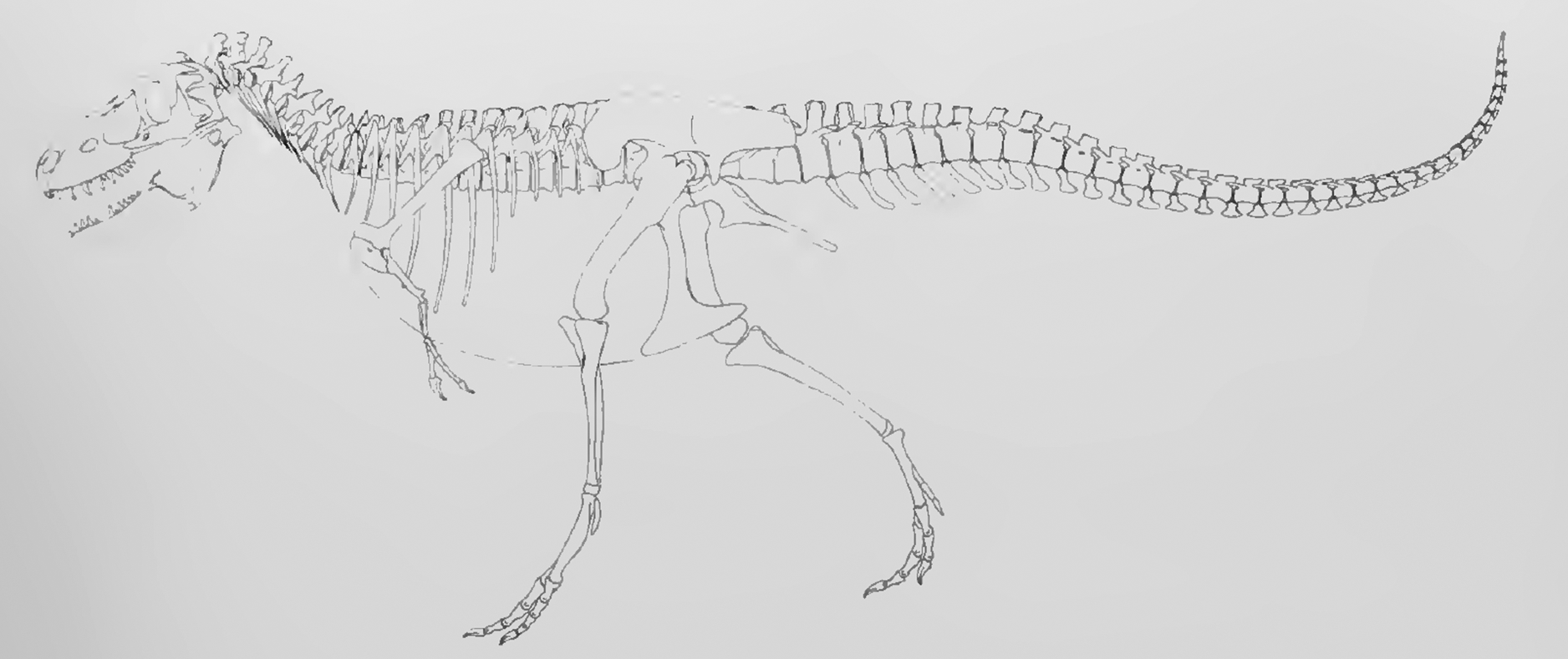
The reconstruction of Daspletosaurus torosus ( Figure 9 View Figure 9 ) is based principally on the type skeleton (NMC 8506) and the hind limb of the paratype ( AMNH 5438 View Materials ). Restored parts include the last two phalanges of the second digit of the manus, acetabular area of the ischium, posterior tip of the pubic boot, all of the metatarsals except the second, the entire pes, and caudal vertebrae 12-14, 18, 19, 21, 22, 26, 28-30, 32, and 36-37. The hind limbs of NMC 350 and NMC 11594 were consulted in drawing elements missing in this region of the type and paratype. Cervical and thoracic ribs are preserved on one side or the other throughout the presacral series of NMC 8506. In the reconstruction they are drawn as seen in lateral aspect, causing the ribs of the posterior thoracic region, which curve away from the vertebrae at a high angle, to appear unusually short. The solid line representing the ventral limit of the body was obtained by projecting the curvature of the anterior thoracic ribs to the midline of the body and by extending this line back to the anteroventral tip of the pubic boot. The abdominal ribs are not shown in the reconstruction.
The dental formula of D. torosus , as indicated by cranial material referred to the species, is: premaxillary teeth 4, maxillary teeth 14 or 15, dentary teeth 15 or 16. There are 11| to 15 serrations per 5 mm on the carinae of the marginal teeth, the serrations being coarsest near the centre of the jaws.
The palate is disarticulated but exceptionally well preserved in NMC 8506 {see Figure 8 View Figure 8 ). The vomers are fused together anteriorly dividing posteriorly into two thin vertical sheets of bone. A vertical anterior ala from the pterygoid broadly underlaps each of these vomerine processes. However, the pterygoids do not meet each other on the midline of the skull, nor do they contact the ventral part of the ascending processes of the palatines (the same relations exist in the palate of A. libratus , see AMNH negative number 37883, taken of USNM 12814). In Albertosaurus {see NMC 5600; Gilmore 1946: 11) and probably also in Daspletosaurus (NMC 8506) the upper edges of the anterior pterygoid alae loosely but widely contact the upper edge of these palatine processes, with the result that each half of the tyrannosaur palate is separated from the opposite half as far anteriorly as the middle of the vomers. The anterodorsal tip of the ascending process of the palatine is firmly applied to the external dorsoposterior corner of the vomer in D. torosus (NMC 8506) and perhaps also in Albertosaurus ( AMNH 5336 View Materials ?; Gilmore 1946: 11).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |