
Thamnodynastes silvai, Trevine & Caicedo-Portilla & Hoogmoed & Thomas & Franco & Montingelli & Osorno-Muñoz & Zaher, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4952.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:F5DAA394-76EE-46FE-B001-299DA42AD825 |
|
DOI |
https://doi.org/10.5281/zenodo.4694847 |
|
persistent identifier |
https://treatment.plazi.org/id/03E287EB-FFF1-FFF0-CAB1-4BF0FC13FE79 |
|
treatment provided by |
Plazi |
|
scientific name |
Thamnodynastes silvai |
| status |
sp. nov. |
Thamnodynastes silvai sp. nov.
( Figs. 3–4 View FIGURE 3 View FIGURE 4 )
Thamnodynastes cf. pallidus Dixon & Soini, 1977: 73 ; Dixon and Soini, 1986: 132.
Thamnodynastes pallidus Bartlett & Bartlett, 2003: 250 , fig. 248.
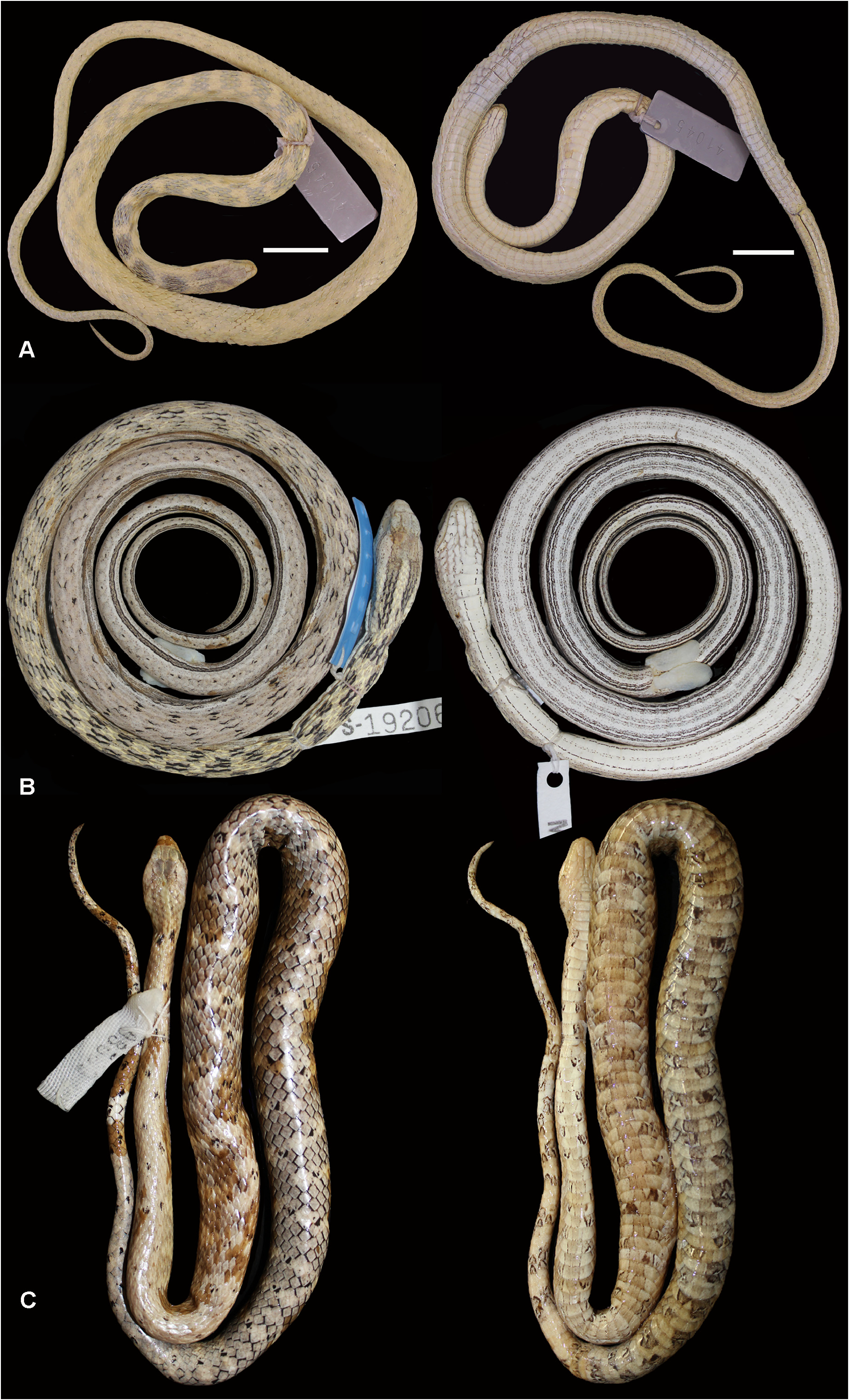
Holotype. An adult male, SINCHI-R 2139 (Field Number JSH 948 ), collected by Juan Silva Haad on Isla Rondinã (no collection date), province of Mariscal Ramón Castilla, district of Ramón Castilla, department of Loreto, Peru ( Fig. 3 View FIGURE 3 ).
Paratypes (n =30). Seven adults and one juvenile from Loreto Department , Peru : female, AMNH 52125 View Materials , collected by Harvey Bassler on March 1931 at Caballococha, Amazon River; male, AMNH 53581 View Materials , collected by Harvey Bassler on December 16 1926 at Rean Rean, Suhuayo (near Contamana , Ucayali River); male , AMNH 52083 View Materials and female , AMNH 55472 View Materials , without additional information; male AMNH 55643 View Materials , collected by Harvey Bassler at Monte Carmelo, Requena Province ( Uresti ), no collection date; female , MHNC 3143 View Materials (Field Number 11) from Dos de Mayo (Sarayacu), Ucayali Province; male , USNM 197292 View Materials (Field Number AJ 933 ), collected on November 1970 at Iquitos ; young male, CORBIDI 6072 View Materials , from Requena Province ( Jenaro Herrera ) without additional information. Twelve adults collected by Dionisio Mafra at Yahuma island , Cayarú , Mariscal Ramón Castilla Province , Ramón Castilla district, Loreto Department, Peru: five females, SINCHI-R 2143 (Field Number JSH 1420 ), SINCHI-R 2154 ( Field Number JSH 1182 ) , SINCHI-R 2148 (Field Number JSH 956 ) , SINCHI-R 2147 (Field Number JSH 1034 ) and SINCHI-R 2151 (Field Number JSH 1585 ); and seven males , SINCHI-R 2140 (Field Number JSH 1051 ) , SINCHI-R 2141 (Field Number JSH 1427 ) , SINCHI-R 2163 (Field Number JSH 1062 ) , SINCHI-R 2146 (Field Number JSH 963 ) , SINCHI-R 2149 (Field Number JSH 704 ) , SINCHI-R 2150 (Field Number JSH 1422 ), and SINCHI-R 2157 (Field Number JSH 1043 ). Five adults and one juvenile from Leticia , Amazonas department, Colombia : female, IAVH 2046 View Materials ( Field Number OPM 518 View Materials ) , collected by Oscar Pinto-Moreno at Amacayacu Natural National Park ; young male, IAVH 854 View Materials , collected by Juan Silva Haad on August 1978; two males, SINCHI-R 2144 (Field Number JSH 1127 ) and SINCHI-R 2169 (Field Number JSH 861 ); and two females , SINCHI-R 2142 (Field Number JSH 819 ) and SINCHI-R 2159 (Field Number JSH 150 ). One adult male , USNM 233065 View Materials (Field Number 807), collected on May 1952 at Maruma, Morona-Santiago Province, Ecuador (no collector information) . Two adult females, SINCHI-R 2153 (Field Number JSH 373 ) and SINCHI-R 2162 (Field Number JSH 429 ), from Tabatinga , Amazonas State , Brazil . One adult female, MPEG 18835 View Materials (Field Number TCAP 2357 ), collected on July 29, 1994 by Marinus S. Hoogmoed and Teresa C. Ávila-Pires at the Mamirauá Ecological Station , Araçazinho lake (left bank of Paraná Apara ), Uarini, Amazonas state, Brazil .
Diagnosis. Venter scattered with sparse dark brown blotches, not forming longitudinal ventral stripes; 12–21 dark brown blotches on the first third of the dorsum of the body, which progressively fade and turn into small brown spots on each side of the paravertebral region ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ); dorsal scale rows smooth, in 17/17/11 (70%, n= 42) or 17/17/13 (22%, n= 13) series; dorsals lacking apical pits; ventrals 124–137 in females and 127–148 in males; and subcaudals 80–97 in females, 83–101 in males ( Table 2).
Comparisons. Thamnodynastes silvai differs from all congeners by having 17/17/13 or 17/17/11 dorsals [vs. 19/19/ 15 in T. almae Franco & Ferreira, 2003 ; T. chaquensis Bergna & Alvarez, 1993 ; T. corocoroensis Gorzula & Ayarzagüena, 1996 ; T. dixoni ; T. gambotensis Perez-Santos & Moreno, 1989 ; T. hypoconia ; T. marahuaquensis Gorzula & Ayarzagüena, 1996 ; T. nattereri ; T. paraguanae ; T. phoenix Franco, Trevine, Montingelli & Zaher, 2017 ; T. ramonriveroi Manzanilla & Sánchez, 2005 ; T. rutilus (Prado, 1942) ; T. strigatus ; T. yavi Myers & Donnelly, 1996 ; and 19/19/ 13 in T. ceibae Bailey & Thomas, 2007 ], and ventral color pattern lacking longitudinal stripes (vs. at least some portion of venter with continuous or inconspicuous longitudinal stripes in all other Thamnodynastes , except for T. sertanejo ). In addition, from the other species with 17 dorsals at midbody ( T. chimanta Roze, 1958 , T. duida and T. lanei Bailey, Thomas & Silva Jr., 2005 ), the new species differs by having 13 series of dorsals on the cloacal region (vs. 15 series in T. chimanta , T. duida and T. lanei ); smooth dorsal scales (vs. dorsals markedly keeled in T. lanei ); and 80–101 subcaudals (vs. 51–61 subcaudals in T. chimanta , 56 in T. duida and 73–88 in T. lanei ) ( Table 1). From the species of the T. pallidus group ( T. longicaudus , T. pallidus and T. sertanejo ), T. silvai differs by a set of characters ( Table 2). Besides the absence of longitudinal ventral stripes, it differs from T. longicaudus by having 17/17/13 or 17/17/11 dorsals (vs. 19/19/13); from T. pallidus by divided cloacal (vs. single), the overall dorsal pattern, composed by several blotches on the first third of the body (vs. first third with several spots of lighter scales, yellow and whitish, bordered by dark brown or black markings in the scale interspaces, in a variegated pattern, sometimes forming smaller blotches), and wider and darker postocular stripe and supralabials spots (vs. postocular stripe and labial spots composed only by a thin dark line delineating the border, not colored internally) ( Figs. 1 View FIGURE 1 , 3 View FIGURE 3 ). Thamnodynastes sertanejo is the only other member of the genus with a blotched ventral pattern (not forming longitudinal stripes). Even so, T. silvai can be easily differentiated by head length 12.3–17.1 mm (vs. 16.1–25.8 mm in T. sertanejo ), lower number of ventral counts, both for male and females ( Table 2), and also by the overall dorsal body pattern: transverse dark brown or greyish bands separated by lighter bands (formed by three to five rows of beige scales) on the first third of the body, progressively vanishing into a homogeneous pattern with a few scattered small dark spots in mid-body that continue throughout the tail (vs. generally homogeneous first third of the body in T. sertanejo , with dark broad dorsal blotches separated by transverse bands of two or three rows of white scales, evident from the mid-body to tail) ( Figs. 1 View FIGURE 1 , 3–4 View FIGURE 3 View FIGURE 4 ). Additionally, T. sertanejo also exhibits a few scattered black dots on the anterior portion of the venter, which may appear as larger black spots on the ventral posterior region ( Fig. 1 View FIGURE 1 ).
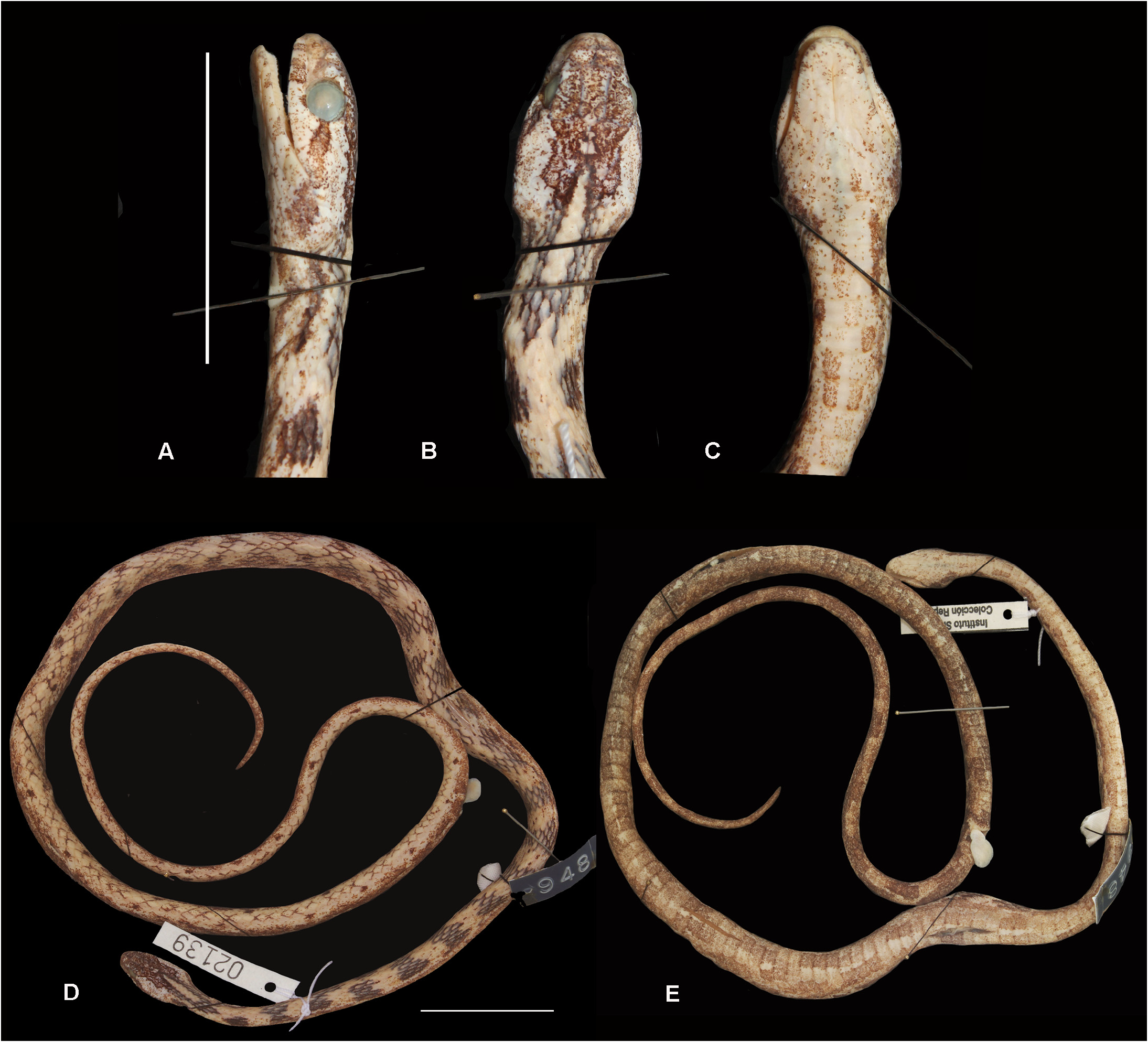
Description of the holotype. Dorsals smooth in 17/17/11 rows; apical pit absent; ventrals 136 and divided subcaudals 98; cloacal semi-divided (division on half of the scale length); supralabials 8/8, 4–5 th contacting eye; infralabials 10/9, 1–5 th /1–4 th contacting first pair of mentals, 5–6 th /4–5 th contacting the second pair; pair of mental shields, approximately the same size; temporals 2+3/2+3; nasal single; loreal elongate on the right side, slightly square on the left; preoculars 1/1 and postoculars 2/2; SVL 440 mm, TL 237.5 mm, head length 15.3 mm, head height 6.1 mm, head width 8.5 mm; distance between nostrils 2.5 mm, eye height 2.6 mm, eye length 3 mm, distance between eyes 5.4 mm, eye-mouth distance (approximately at the level of the fifth supralabial) 1.2 mm, eye-rostral distance 3.6 mm; internasal scale (length X width): 1.2 X 0.9 mm, prefrontal: 1.7 X 1.5 mm, frontal: 4.4 X 1.7 mm, parietal: 4.5 X 2.2 mm, loreal: 1.1 X 0.6 mm; prediastemal maxillary teeth 9 plus 2 slightly enlarged and grooved fangs.
In preservative, dorsal surface of the head with intense brown pigmentation from the supratemporals to internasals, temporals light beige with several small brown dots; brown postocular stripe, extending from the postoculars to the rear corner of the mouth, where it continues as a blotch of seven scales in length; supralabial background light beige with a few small brown dots, brown blotches on the first three supralabials, and a small blotch right under the eye between supralabials four and five; infralabials background light beige with scattered small brown stains, reaching mentals, gulars and preventrals. Sixteen dorsal dark brown bands, two to four scale rows long, more prominent on the first third of the body, progressively losing pigmentation towards the tail, until fading into sparse dark brown spots on each side of the vertebral scale; beige stripe from the distal border of the parietals, extending eight scales onto the neck, bordered by two dark brown stripes (10 scales long) on each side; ventral background light beige with intense pigmentation of minute brown spots, darkening the overall ventral pattern towards the tail; diffuse dark brown blotches, concentrated on the paraventral portion, more evident from the second third of the body; ventral portion of the tail dark brown with more intense pigmentation than body ( Fig. 3 View FIGURE 3 ).
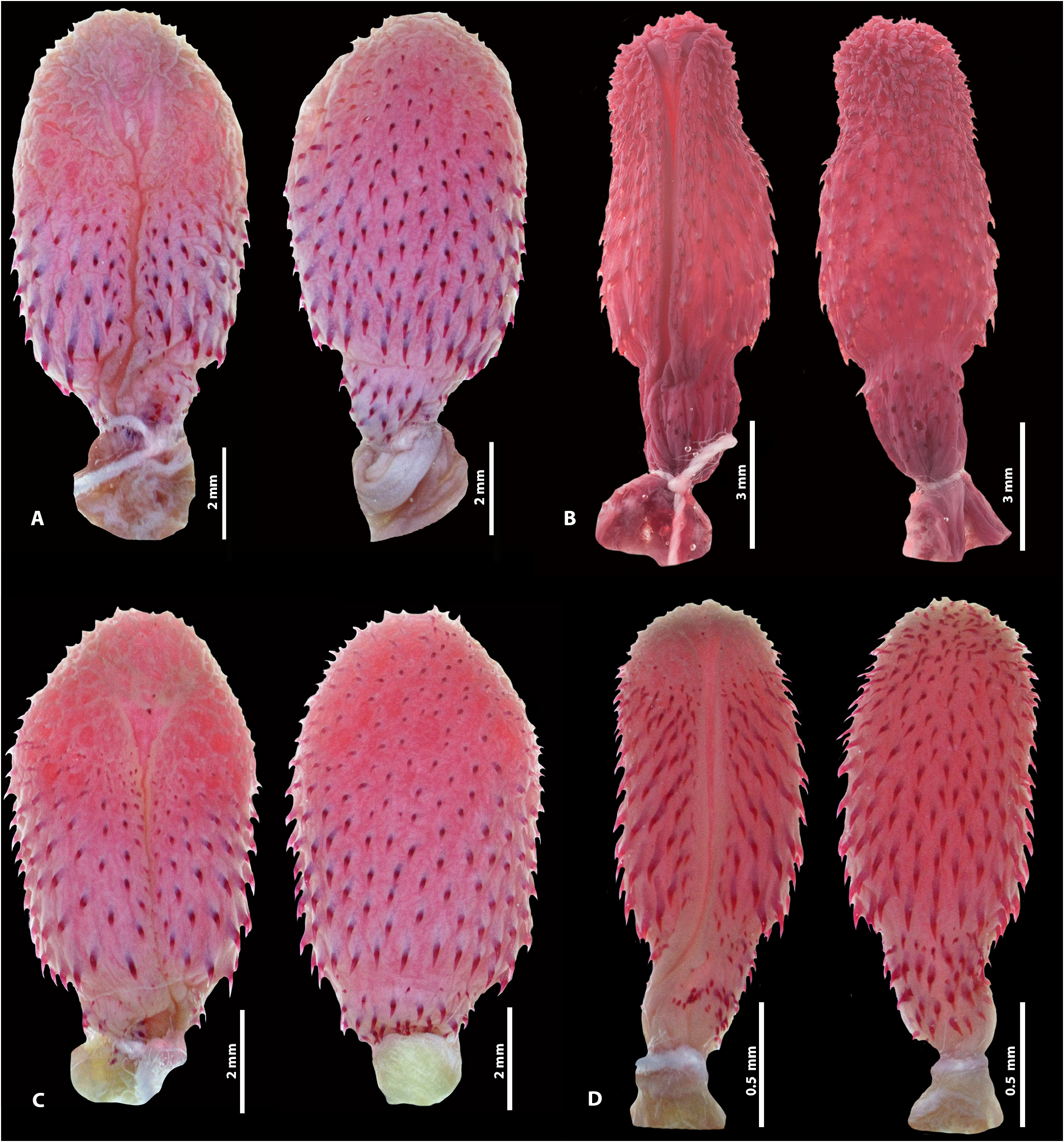
The hemipenis of the holotype is unilobed (although with divided retractor muscle), unicalyculate and non-capitated, with very short capitulum covered with small papillate calyces; few spinulate calyces on the proximal portion of capitulum on the asulcate side; hemipenial body covered with a few rows of small spines, 19 slightly larger spines encircling first row; sulcus spermaticus divided at the base of the capitulum, with sulci ending in slightly enlarged area, on approximately half the capitulum length; subtle constriction between base and hemipenial body; base covered with several spines, approximately the same size as the body spines ( Fig. 5 View FIGURE 5 ).
Hemipenial morphology (n =15). Thamnodynastes silvai has a hemipenis pattern corresponding fairly well to the overall pattern of the T. pallidus group, more closely resembling the hemipenis of T. longicaudus ( Figs. 2 View FIGURE 2 , 5 View FIGURE 5 ). Additional prepared hemipenes exhibited little variation ( Fig. 5 View FIGURE 5 ), such as: capitular spinulate calyces can be more evident on the proximal portion of the capitulum, on the asulcate side ( Fig. 5C View FIGURE 5 ); the sulci are short, after the sulcus spermaticus division, associated with a short capitulum in all analyzed specimens, and their termination can be slightly closer to the capitulum apex ( Fig. 5B–D View FIGURE 5 ).
Meristic and morphometric variation. Largest male (SINCHI-R 2139) SVL 440 mm and TL 237.5 mm; largest female (AMNH 55472) SVL 388 mm and TL 170 mm; smooth dorsal scales in 17/17/11 (70%, n =42), 17/17/13 (22%, n =13), 15/17/11 (3.3%, n =2), 17/17/12 (3.3%, n =2) or 17/17/10 (1.6%, n =1) series. Sexual dimorphism was reported for both ventrals (t = -8.4707, p<0.001) and subcaudals (t= -6.0197, p <0.001); ventrals 127–148 (137±4.8, n =27) and subcaudals 83–101 (94± 4.2, n =21) in males, ventrals 124–137 (128±2.9, n =31) and subcaudals 80–97 (87±4.1, n =28) in females; supralabials 8 (98%, n =52) or 8/7 (2%, n =1); 4–5 th supralabials contacting eye (75%, n =38) or 3– 5 th (28%, n =15); infralabials 9 (75%, n =43), 8/8 (8.7%, n =5), 9/8 (5.3%, n =3), 9/10 (3.5%, n =2), 10/9 (3.5%, n =2), 8/9 (1.7%, n =1) or 10/10 (1.7%, n =1); preoculars 1/1 (93%, n =52) or 1/2 (n =1); postoculars 2/2 (86%, n =49), 2/3 (n =1), 3/2 (n =2), or 3/3 (n =2); nasal scale single (98%, n =52) or semi-divided on the right side (n =1).
Color pattern variation. In preservative, examined specimens show head usually very pigmented with several small dots, from rostral to temporal scales, almost black in few specimens (USNM 233065, MHNC 3143). Head usually with two dark brown bands right after temporals, extending onto the neck, occasionally merged with first pair of dorsal blotches (USNM 203065, SINCHI-R 2142, 2139) ( Fig. 4 View FIGURE 4 ). These two bands may define a lighter central neck band (SINCHI-R 2153). Postocular stripe very distinguished, the same color as dorsal blotches, ending right after the corner of the mouth, and extending for 2 or 3 scales ventrally (IAVH 2046), or linearly extending on the side of the body (AMNH 52083) ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ). Venter of the head light beige, with a few scattered brown dots concentrated more on neck region and laterally. Twelve to 21 dorsal bands on the first third of the body, defined into regular transverse bands (CORBIDI 6072, AMNH 52536) or irregularly shaped (e.g. IAVH 2046) ( Fig. 4 View FIGURE 4 ). Final dorsal two thirds of body and tail with more intensely pigmented variegate pattern (AMNH 52083, 53581), or with very subtle pigmentation, almost immaculate (AMNH 52858). All specimens (n =64) lack longitudinal ventral stripes, with ventral brown blotches dispersed throughout the belly and tail ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ). These blotches can be evenly spaced, occupying two ventral scales each (AMNH 53582, 55472), or more irregularly dispersed (IAVH 2046, USNM 233065) ( Fig. 4 View FIGURE 4 ). Most specimens with overall ventral darkening towards the tail, from beige or light cream on the head to an intensely pigmentated brown tail (IAVH 2046).
Color in life. The following pattern and coloration description of specimen AMNH 147707, from Pebas, Loreto Department, Peru, is from the field notes of Charles W. Myers (pers. comm. to Robert A. Thomas dated April 14, 1977): “Overall grayish brown above, with a few blackish brown spots and vague yellowish-brown areas that are paler than ground color and that occur at intervals to give an incipient banded pattern. The neck or hood region is banded with brown saddles and lighter brown interspaces (scales in the interspaces have pale yellow bases set in white skin). A pair of vague dark brown lines on rear of head and nape. Underside head and neck white, thence turning an increasingly dark grayish brown on venter, with squarish marks of still darker brown; these marks tend to be broken and/or bordered by streaks of whitish. Iris pale brown; pupil vertical. Tongue overall grayish black except for a broad brown band that encircles the base of fork.”
Etymology. The specific epithet “ silvai ” is patronymic honoring the late Juan Silva Haad, physician at the hospital of Leticia for many years, who maintained a snake collection for over 50 years. He collected several species from Colombian, Peruvian and Brazilian Amazonia, including several of the Thamnodynastes specimens used in this description. He donated most part of his collection to the Sinchi Institute, with over 2,500 specimens of Amazonian snakes, which represented an immense contribution for the Institute’s collection, and a valuable addition of important data for the study of Amazonian snakes. Unfortunately, he passed away on January 31, 2021, just a few weeks before the final reviewed version of the manuscript was accepted.
Distribution and natural history. Six females had three to six developed follicles with visible embryos (possibly in early stages of development due to their small size): SINCHI-R 2142 (3 follicles/embryos), SINCHI-R 2143 (6), SINCHI-R 2147 (4), SINCHI-R 2154 (5), SINCHI-R 2172 (5), and SINCHI-R 2173 (5); and one female (MPEG 18835) had three well-developed follicles with larger visible embryos, confirming a viviparous reproductive mode. The largest pregnant female (SINCHI-R 2147: SVL 323 mm, TL 173 mm) had four embryos. One specimen (AMNH 53582: SVL 321 mm, TL 152 mm) had a small frog (family Hylidae ) in the stomach.
On July 29, 1994 at 14:15 h, during the flood season in the Amazon, Marinus S. Hoogmoed and Teresa C. Avila-Pires collected a female ( MPEG 18835 View Materials ) in an isolated tree, emerging from a floating meadow (mixed vegetation) at the edge of a lake, completely surrounded by floating grassy mats along its banks. The specimen was slowly moving on the branches of a tree at a height of about 2m above the water surface. When collected (and later again when photographed), it flattened its head and anterior part of the body horizontally (forming a hood), which caused the thin skin between the scales to be stretched and exposed, suddenly showing a bold pattern of transverse, dark brown spots on a lighter background ( Figs. 6A–B View FIGURE 6 ). Within the spots, the edges of the scales were black, forming three to six horizontal short zig-zag lines (about three scales long) within the dark spots. Because of the abrupt change this behavior was interpreted as being defensive. No attempt to bite was made. We also report, for the first time, the same behavior for T. pallidus . Two specimens ( SINCHI-R 2528 and 2529) displayed the hood-displaying performance while their photographs were being taken, during fieldwork in San José del Guaviare, Guaviare Department, Colombia, in June 15, 2017 ( Figs. 6C–D View FIGURE 6 ) .
Thamnodynastes silvai occurs in forested areas of Amazonia, along the triple frontier of Colombia, Brazil and Peru; extending southwest to the Loreto Department in Peru, northwest to the province of Morona-Santiago in Ecuador, and northeast to Uarini, state of Amazonas in Brazil ( Fig. 7 View FIGURE 7 ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Thamnodynastes silvai
| Trevine, Vivian C., Caicedo-Portilla, José Rancés, Hoogmoed, Marinus, Thomas, Robert A., Franco, Francisco L., Montingelli, Giovanna G., Osorno-Muñoz, Mariela & Zaher, Hussam 2021 |
Thamnodynastes pallidus
| Bartlett, R. D. & Bartlett, P. 2003: 250 |
Thamnodynastes cf. pallidus
| Dixon, J. R. & Soini, P. 1986: 132 |
| Dixon, J. R. & Soini, P. 1977: 73 |