
Loxosoma axisadversum Konno, 1972
|
publication ID |
https://doi.org/10.12782/sd.20.1.083 |
|
persistent identifier |
https://treatment.plazi.org/id/03E1879D-1C11-441A-FD6F-F9F6FBC0E87A |
|
treatment provided by |
Felipe |
|
scientific name |
Loxosoma axisadversum Konno, 1972 |
| status |
|
Loxosoma axisadversum Konno, 1972 View in CoL
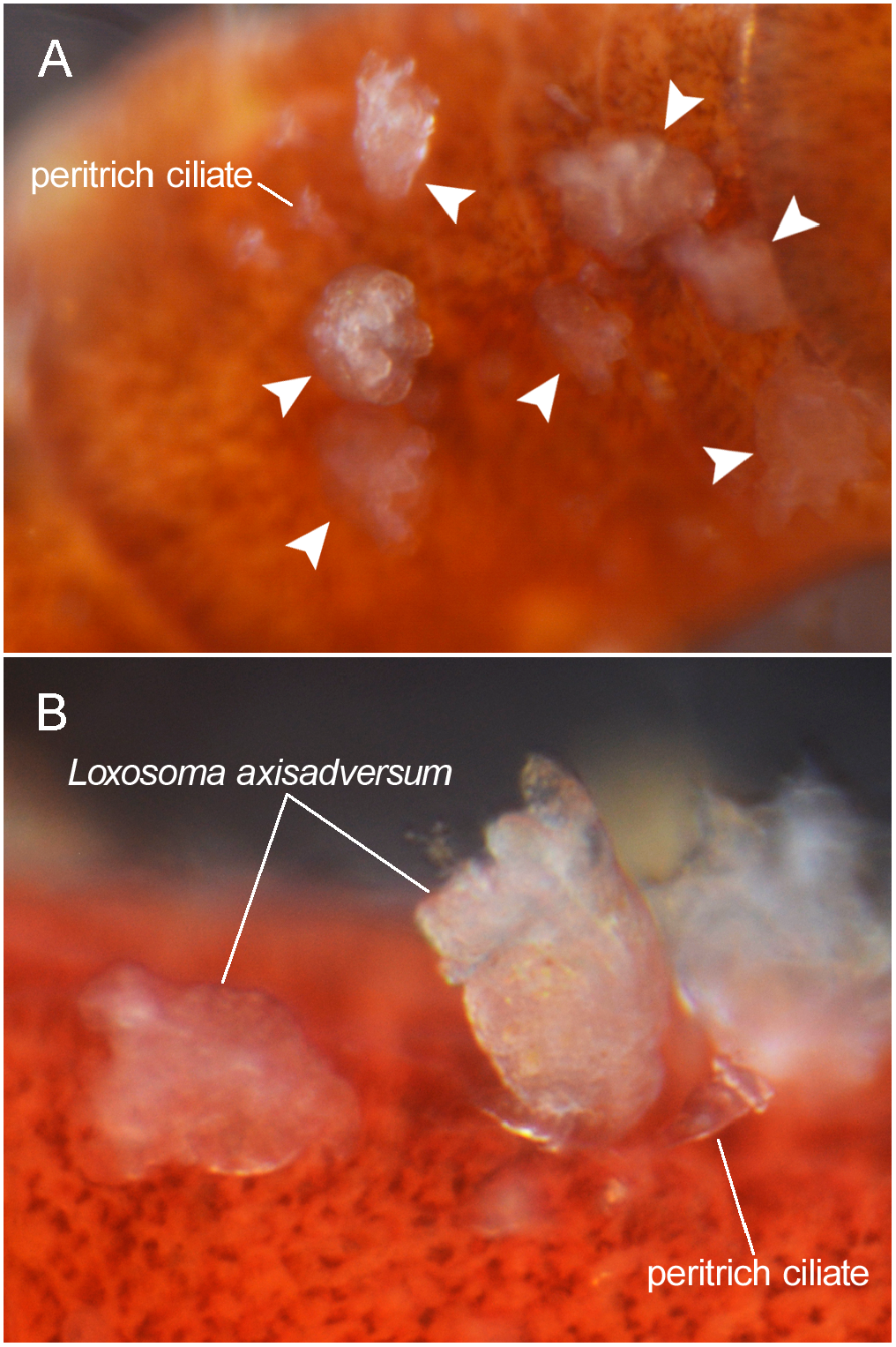
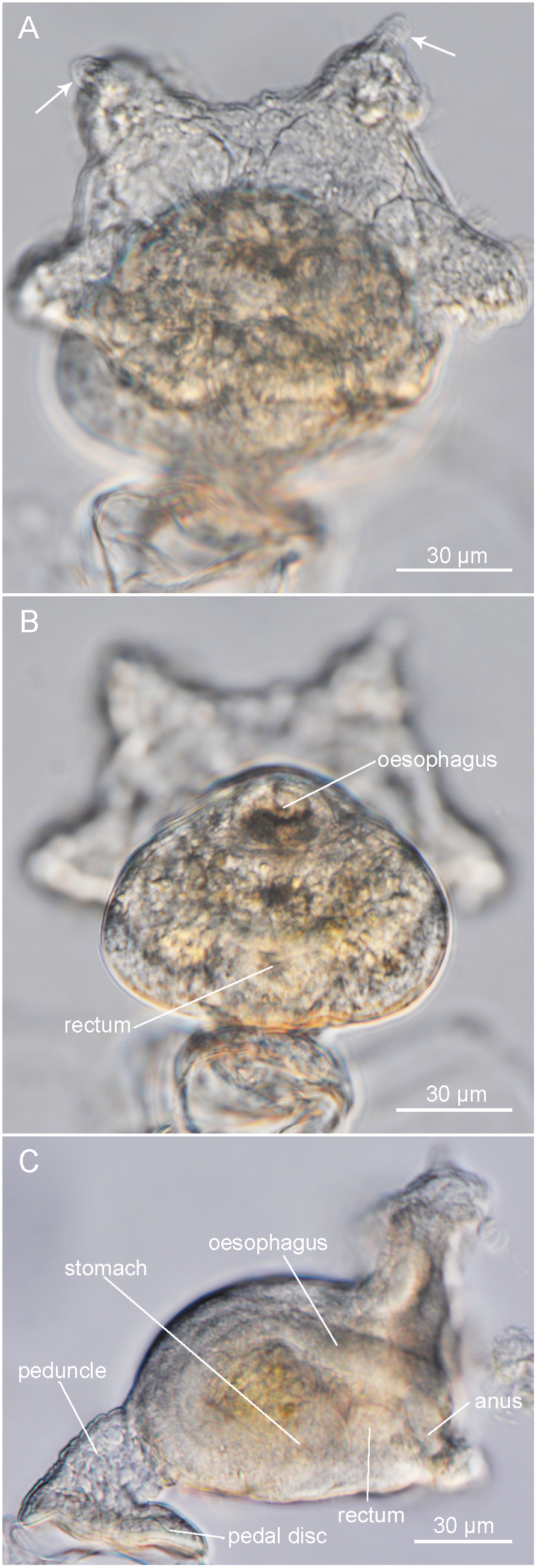
[Japanese name: Ushiromae-rokusosoma] ( Figs 2 View Fig , 3 View Fig )
Loxosoma axisadversum Konno, 1972: 23–24 View in CoL , figs 1–8; Konno 1978: 22–23, figs 9–11; Konno 1985: 5.
Material examined. Ten or more specimens alive, of which one was observed under compound microscope: ZIHU 4935 View Materials ( five specimens fixed in 10% formalin–seawater, preserved in 90% EtOH), ZIHU 4936 View Materials (a few individuals attached to the host polychaete, fixed and preserved in 10% formalin–seawater), and five specimens destroyed for DNA extraction .
Description of one specimen. Destroyed for DNA extraction after observation. Body translucent ( Fig. 2A, B View Fig ). Total body length 150 µm. Tentacle crown 140 µm in diameter, with three pairs of very short tentacles ( Fig. 3A View Fig ); those of two larger, upper pairs each with small papilla on distal, abfrontal side near tip ( Fig. 3A View Fig ). Mouth situated above anus within tentacle crown ( Fig. 3B View Fig ). Calyx oval in shape, 90 µm in length and 100 µm in diameter. Peduncle stout, 30 µm long and 25 µm wide ( Fig. 3B, C View Fig ). Pedal disc rounded, 60 µm in length and 40 µm in width.
Distribution on host. Found most densely on dorsal side slightly anterior to middle portion of polychaete’s body, with up to about eight individuals on chaetiger 5 (or 6), other individuals mostly found back to chaetiger 10.
Host species. Nicomache minor ( Konno 1972, 1978, 1985); N. personata (this study).
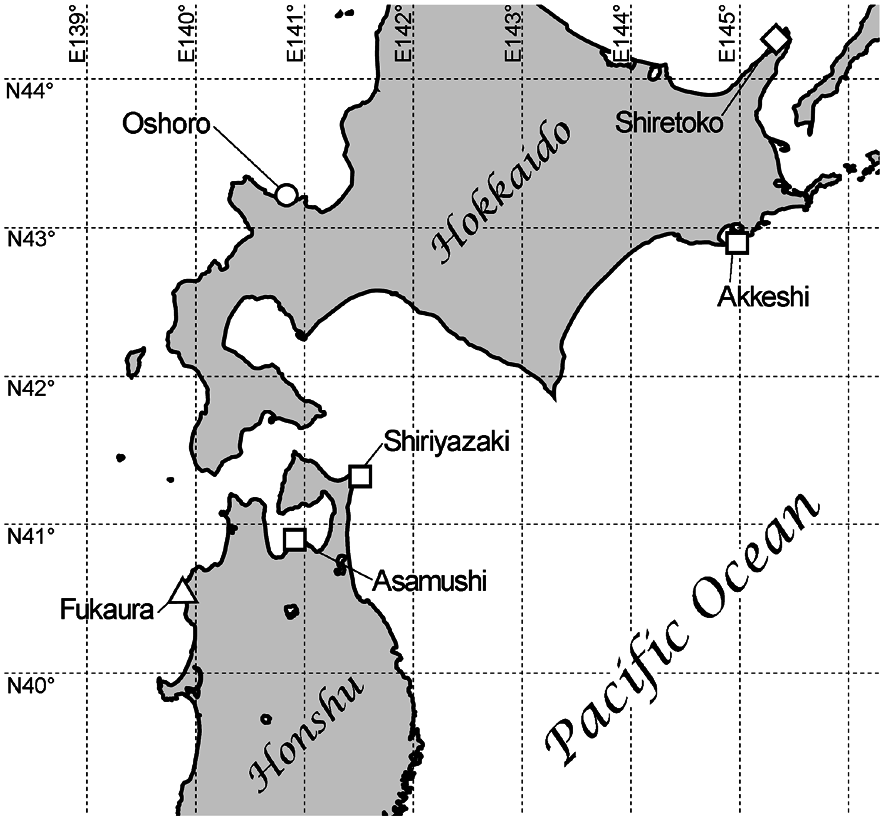
Remarks. The character of the present material agrees well with the morphological account given in the original description by Konno (1972). The body size of the present material falls within the reported range of the species in the original description ( Konno 1972). The close geographical proximity of the present sampling site (Oshoro) to the type locality (Fukaura) ( ca. 300 km; Fig. 1 View Fig ), both under the influence of the Tsushima Current, further justifies our identifi- cation of the material.
Loxosoma axisadversum has previously been known only in northern Japan from one host species, Nicomache minor (Annelida: Maldanidae ). The latter has been reported to be widely distributed in cold waters in the Northern Hemisphere: Norwegian Sea ( Arwidsson 1907), Barents Sea ( Arwidsson 1907; Kędra et al. 2010), White Sea ( Tzetlin and Markelova 1985; Tzetlin et al. 1997; Solyanko et al. 2011), Sea of Japan ( Annenkova 1937; Ushakov 1955; Imajima and Shiraki 1982; Yakovlevich 2013), Sea of Okhotsk ( Ushakov 1955; Imajima and Shiraki 1982), Pacific coasts of northern Japan ( Imajima and Shiraki 1982), Bering Sea ( Ushakov 1955), and Hudson Bay ( Berkeley and Berkeley 1943). In the White Sea, Tzetlin and Markelova (1985) found 15 species of symbionts in the tubes of N. minor , of which only an unidentified species of “ Loxosomella ” appeared to possess a species-specific relation with the polychaetes. Unfortunately, Tzetlin and Markelova (1985) did not provide any further information, so the identity of their material remains unknown. More recently, however, Borisanova and Krylova (2014) summarized 10 species of Loxosomella reported from the White Sea, of which L. polita Nielsen, 1964 and L. similis Nielsen, 1964 are associated with N. minor (Borisanova in litt.), but no species of Loxosoma has hitherto been reported from this sea area.
We identified the present host maldanid as N. personata because it possessed 22 chaetigers and the anal funnel was equipped with 16 papillae ( Johnson 1901; Imajima and Shiraki 1982; Imajima 1996; De Assis et al. 2007); ours is the first record of L. axisadversum from this host species. However, the two maldanids N. minor and N. personata are quite similar to each other in shape, and the alleged morphological differences between the two ( i.e., number of chaetigers: 22 in N. personata vs 23 in N. minor ; Khlebovich 1961; Tarakanova 1974; Imajima and Shiraki 1982) may simply represent intraspecific variation; indeed, they were once regarded as conspecific ( Pettibone 1954). A BLAST search ( Altschul et al. 1997) at the NCBI website (http://blast.ncbi.nlm.nih. gov) with the 331 nt of the histone H3 sequence of what we identified as N. personata from Oshoro (DDBJ accession number: LC005496 View Materials ) showed that it was 97% identical (with 93% query coverage) to KF926672 View Materials , a sequence derived from a polychaete identified as N. minor collected in the White Sea (T. Neretina et al., unpublished). The two sequences differ by 1.8% in terms of uncorrected p -distance, indicating that the two populations—more than 6,000 km away from each other—are at least mutually closely related, if not the same species. In other annelids, maximum intraspecific difference at the same locality with regard to histone H3 sequence was 1.1% in the opheliid Ophelina cryptophila Neave and Glasby, 2013 from Darwin Harbour (between JN182668 View Materials and JN182673 View Materials ; Neave and Glasby 2013), while the minimum interspecific difference between O. cryptophila and O. tessellata Neave and Glasby, 2013 was 3.3% ( e.g., between JN182680 View Materials and JN182667 View Materials ; Neave and Glasby 2013). Nicomache minor and N. personata together show a circumpolar distribution but co-occur in Japanese waters ( Imajima and Shiraki 1982; De Assis et al. 2007), suggesting the possibil- ity of a ring-species complex ( Alcaide et al. 2014), although resolving this point is beyond the scope of this paper.
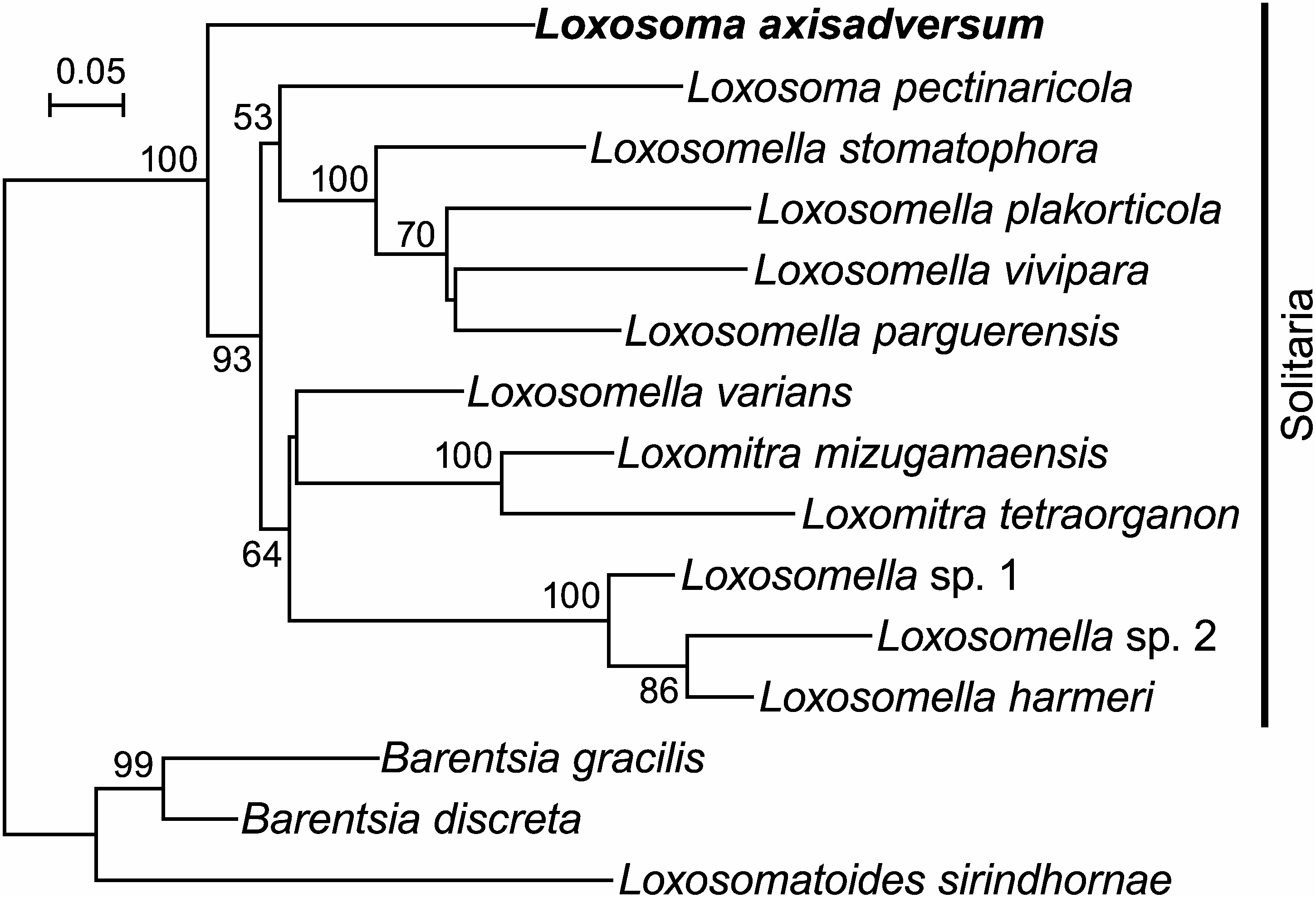
Phylogeny. Of the five specimens from which we extracted DNA, PCR amplification of all the three gene markers was successful in only one (specimen number 4, Table 1). The sequences from this specimen were included in the analysis. The other determined sequences from different specimens were identical with respect to each gene marker, except for a single, synonymous substitution in COI. In the resulting ML tree based on the concatenated COI+18S+28S dataset ( Fig. 4 View Fig ), L. axisadversum appears as sister to all the rest of the solitarians included in the analysis. The same topology was observed in ML trees based only on 18S or 28S, while the COI tree was not well resolved (data not shown). The diagnosis and circumscription of the genus Loxosoma based on foot morphology ( Mortensen 1911; Nielsen 1996) thus do not appear to reflect the evolutionary relationships among solitary entoprocts.
That L. axisadversum and L. pectinaricola turned out not to be sister taxa in our analysis may reflect the difference in mode of attachment of their buds to the parent. Based on this feature, Nielsen (1964) proposed a subgeneric discrimination within Loxosoma , placing species with buds attached to their parents by the abfrontal margin of the sucking disc in the subgenus Loxomorpha Nielsen, 1964 (preoccupied and renamed as Loxosomina Nielsen, 1996), while species with buds attached by the central area of the sucking disc were classified into the nominotypical subgenus Loxosoma . Loxosoma axisadversum and L. pectinaricola exhibit budattachment modes that are typical of Loxosoma ( Loxosomina) and Loxosoma ( Loxosoma) , respectively ( Nielsen 1964, 1996; Konno 1972), and this suggests that reassignment of L. axisadversum to Loxosomina may be in order. Ideally, future analyses with expanded taxon sampling including the type species of all the genus-group names in the family Loxosomatidae should be done to arrive at a better taxonomy of this group with proper application of the genus names reflecting their phylogeny.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Loxosoma axisadversum Konno, 1972
| Kajihara, Hiroshi, Tomioka, Shinri, Kakui, Keiichi & Iseto, Tohru 2015 |
Loxosoma axisadversum
| Konno, K. 1985: 5 |
| Konno, K. 1978: 22 |
| Konno, K. 1972: 24 |