
Eomys, Schlosser, 1884
|
publication ID |
https://doi.org/ 10.5252/g2010n2a3 |
|
persistent identifier |
https://treatment.plazi.org/id/03E18794-0F17-7244-CEB0-FF75FF1CFD00 |
|
treatment provided by |
Marcus |
|
scientific name |
Eomys |
| status |
|
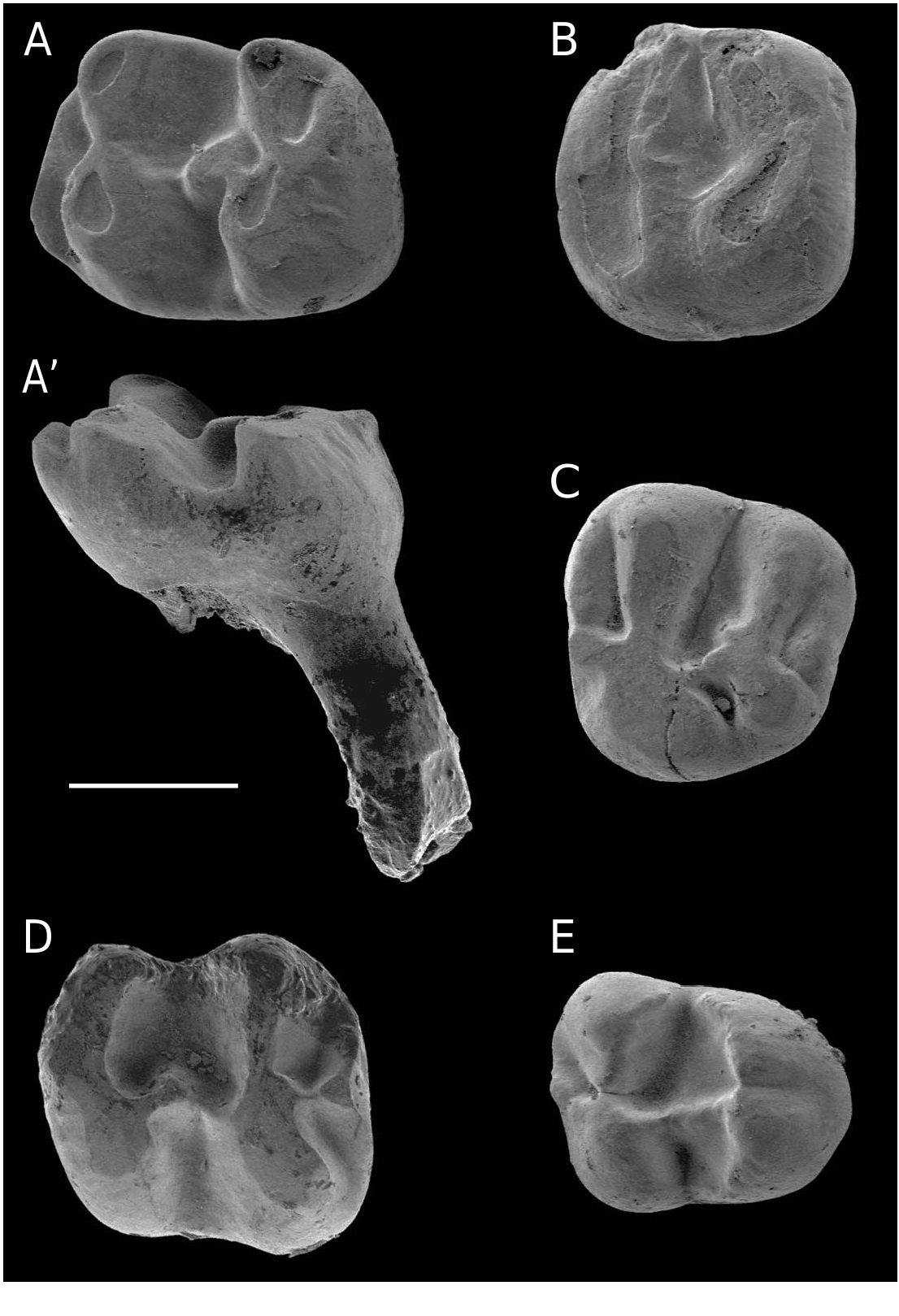
Eomys aff. E. antiquus ( Aymard, 1853) ( Fig. 5 View FIG A-Q)
LOCALITIES. — French and Spanish localities: Saint- Martin-de-Castillon C and F ( France); Montalban ( Spain).
TEETH SIZE. — At a first approach of the material, species can be differentiated on size. We first describe the species characterized by small sized teeth found in Saint-Martin-de-Castillon C, F and Montalban, the size of which also corresponds to that of the smaller sized German species. Measurements for Saint-Martin-de- Castillon levels and Montalban are given in Tables 4-6 and Figure 7 View FIG . Th is species is clearly differentiated by its size compared to the very small sized species found in La Blache and the medium-sized species found in Saint- Martin-de-Castillon J and E.
DESCRIPTION
P4
The general morphology is similar to that of the German material, but some differences can be observed on the labial anteroloph development. The labial anteroloph is generally absent or weakly developed, more rarely well developed.Th e lingual anteroloph is not clearly developed, usually limited to a weak fold of the enamel suggesting the formation of a crest.
The mesoloph has a noticeable variability, it can be absent, weak or short.When short, the mesoloph can be perpendicular to the axis of the tooth or oblique to the front of the tooth. Th ree teeth in Saint-Martinde-Castillon present a discontinuous mesoloph. The rest of the teeth have the same type of morphological variability as in the German material.
The entoloph is usually continuous, but can sometimes be broken between the mesoloph and the protocone or between the mesoloph and the hypocone.
M1/2
On the anterior part of the tooth the labial anteroloph is long, starting at the base of the protoloph. Few teeth have a lingual anteroloph, but when present it can be either weakly or clearly developed. Some rare teeth can have the two anterolophs disconnected from the protoloph. Th e mesoloph is generally short and curved backward, but on some teeth it can be oblique to the front of the tooth. The mesoloph can also be weak, absent, or long and interrupted in some rare cases at Saint-Martin-de-Castillon.
Abbreviations: N, number of teeth; Min, minimal value; Max, maximal value; Mean, mean value; SD, standard deviation; CV, coefficient of variation.
Some teeth clearly present two mesolophs (Appendix Fig. A2-I View FIG ).
As for the P4, the entoloph is usually continuous, but can sometimes be broken between the mesoloph and the protocone or between the mesoloph and the hypocone, or even both at the same time, leading to an isolated mesoloph (Appendix Fig. A2 View FIG -Rare morphologies). One tooth in Saint-Martin-de-Castillon has a paracone disconnected from the entoloph (Appendix Fig. A2 View FIG -Rare morphologies).
M3
The general shape of the tooth is triangular round- ed, its posterior part is generally larger than in the German material. Th e anterior part of the tooth has little morphological variability. The labial anteroloph is always long, starting from the base of the paraloph. A short and weaklydeveloped lingual anteroloph can be observed on some teeth.
The morphology generally has a higher variability on the posterior part around the metaloph. The metaloph is usually completely developed, reaching the labial border, but it can also be short or absent.
A forward crest (mesoloph) can start from the metaloph. It can be short or reaching the labial border of the tooth, and in some rare cases connected to the protoloph (Appendix Fig. A3 -Rare morphologies). A backward crest can also appear, linking the metaloph and the posteroloph.
p4
The general morphology is similar to that of the material from the German localities but some slight differences can be observed in the variability. The anterior part is always divided into two conids. Most of the time they are linked by a transverse crest, but they can also be almost joining each other or both linked backward on the ectolophid, but are never clearly separated. An anteroconid, connected to the protoconid, can sometimes be observed and for one tooth this cusp takes the shape of a real anterolophid (one case observed in Montalban).
The mesolophid is generally short but with a noticeable variability, it can be absent, weak or reaching the lingual border. In some rare cases, the mesolophid can be directly connected to the metaconid by a fore-spur (one case observed in Saint-Martin-de-Castillon C). In the posterior part, the posterolophid is often weakly developed, closely positioned to the hypolophid, delimiting a narrow posterosinus, but it can also be well developed. Some teeth do not show a posterolophid. The ectolophid is generally continuous but some teeth have a disconnection between the protoconid and the mesolophid ( Fig. 5K View FIG ).
m1/2
In the anterior part of the tooth, the labial and lingual anterolophids are generally both well developed, but the labial one can sometimes be shorter or even absent. When the labial anterolophid is strongly developed, the sinus between it and the protoconid can be very wide leading to a round shape of the anterolophid that is clearly observable in the tooth outline. Th e protoconid is generally oriented forward ( Fig. 5 View FIG N-P).
The anterolophids are usually connected to the protoconid but the connection can sometimes be more lingual, being connected to the metalophid, and sometimes disconnected, leading to an isolated anterior crest. The mesolophid has little variability, generally of average size, very rarely weak ( Fig. 5O View FIG ) or absent. Th e ectolophid can sometimes be interrupted leading to an isolated mesolophid in the middle of the tooth. Th e posterolophid is always present even if sometimes reduced to a spur.
m3
As for the m1/2, the labial and lingual anterolophids are generally both well developed with some variability in their connection that can be with the protoconid or with the metalophid. The anterolophids can also very often be disconnected, leading so to an isolated fore-crest.
The mesolophid is always well developed, of medium size or long, reaching the lingual side of the tooth in most cases. Th e ectolophid is usually continuous, but it can be broken between the mesolophid and the hypoconid.
The entoconid is generally weakly developed, matching the thickness of the posterior crest on its lingual part, but rarely absent.
On some rare teeth, a longitudinal spur can appear between the posterolophid and the mesolophid, starting from the mesolophid or from the posterolophid. Th is spur can be weakly developed or well-developed linking the mesolophid and the posterolophid (Appendix Fig. A6 View FIG -IV).
DP4 and dp4
Some teeth have been found in the French and Spanish localities that have the same morphology as in Möhren 13 and 20.
CONCLUSION ON THE EOMYS AFF. E. ANTIQUUS FRENCH AND SPANISH MATERIAL
For Saint-Martin-de-Castillon C and F eomyids seem to belong to a single population not very different of that of the slightly older locality at Montalban. Considering that morphological variability beyond the type specimen (from Ronzon MP 21) is unknown, the specimens from Montalban (MP 23) and Saint- Martin-de-Castillon (MP 24) are here mainly compared with the specimens from Germany (Möhren 13 and 20, MP 21). Some significant morphological differences can be observed, such as:
– an isolated labial anteroloph in P 4 in Germany (not in France or Spain), the labial anteroloph of P4 is less developed on French and Spanish material and the mesoloph more frequently developed on French material (Appendix Fig. A1 View FIG -III);
– on M1-2, 3-6% of the French teeth show a long interrupted mesoloph that is never seen on German and Spanish material (Appendix Fig. A2-I View FIG );
– the absence of metaloph in M 3 in Germany (not in France or Spain); the metaloph and the mesoloph of M3 are more developed on French specimens (Appendix Fig. A3);
– in German material, a clearly divided anteroconid exists on p4, (not in France or Spain) the connection between the metaconid and the protoconid is rare whereas it is more common in French and Spanish material and the mesolophid seems shorter in German localities (Appendix Fig. A4);
– long mesolophids never exist on m1-2 (Appendix Fig. A5 View FIG );
– the mesolophid is more developed on m 3 in French and Spanish specimens (Appendix Fig. A6 View FIG ).
Concerning the size of teeth ( Fig. 7 View FIG ), a comparison between these localities has been made using the Mann-Whitney test. Th is non-parametric test has been chosen because the discrepancy in the number of specimens between the localities does not allow us to check if the values in each sample are normally distributed. Th e results (see Table 7) confirm the size similarity between Möhren 20 and 13, but also the close size relationship between Saint-Martin-de- Castillon C and F. However a significant difference is observed on the length of the m1/2 between the two levels of Saint-Martin-de-Castillon, the teeth of the upper level, Saint-Martin-de-Castillon F, being larger. Concerning Montalban, the results are more diffi cult to interpret because of the generally low number of specimens, likely to affect the result of the test. Montalban presents generally few significant differences with both the German and French localities. However the M1/2 seem to be shorter than the specimens from both Germany and France which might indicate a geographical differentiation. For the remaining teeth, the size in Montalban appears to be intermediate between the German and French localities. The comparison of Möhren 13 with Saint-Martin-de-Castillon C (both with an important number of specimens) indicates a clear difference in the size on the premolars (the German ones being significantly smaller) and also in the width of M3 and m1/2, which is in accordance with the observations made of the morphology. Th ese results indicate a faint size enlargement over time which is not always the case in later eomyids.
Considering that the localities from Southern Germany are probably much older (more or less 3 Ma) than the French and Spanish localities studied, the differences observed above are likely the result of morphological evolution that more strongly affect the teeth at the extremities of the tooth row. The size and morphology of the type material of E. antiquus (2 m 1/2: 0.85 × 0,85 mm; 0.96 × 0.95 mm) are indeed included in the variability described for Montalban and Saint-Martin-de-Castillon C and F. For this reason the populations of the French and Spanish localities are attributed to Eomys aff. E. antiquus .
DISCUSSION
The general morphology described above for the German, French and Spanish small-sized species is characterized by very brachyodont teeth with a bunodont tooth topology based on four main cusps. The longitudinal crest, rarely interrupted, is more or less lingually-located on upper premolars and molars, and labially-located on lower premolars and molars leading to very long transverse crests (protoloph and metaloph for upper teeth and metalophid and hypolophid for lower teeth) beyond the middle axis of the teeth.
The single mandible found in the type locality does not allow us to address the question of the morphological evolution of E. antiquus mandible between MP 21 and MP 24. However the significant number of teeth found in Möhren 13, Möhren 20 and Saint-Martin-de-Castillon (mainly the level C) allows a more precise morphological comparison in order to characterize such a potential evolution. Indeed, the comparisons show, at first, a very similar morphology, but also point out some differences mainly on premolars and third molars, whereas first and second molars are almost identical.
Among the small Eomys forms known from the Early Oligocene, the particular morphologies described from Lovagny (MP 23) by Engesser (1990) can be found in Möhren 13 as well as in Montalban or Saint- Martin-de-Castillon C and, in our opinion, the teeth of Lovagny can be included in E. antiquus .
From the Early/late Oligocene transition (MP 25 level), the material of Bumbach 1(MP 25: Engesser 1990) has been compared (on cast) with the German and French material. It appears that the morphological features of E. nov. sp. 2 are included in the morphological variability of E. antiquus from the new material of Germany. However, the upper teeth seem to show long mesolophs more frequently. Eomys molassicus Engesser, 1987 is a later form (Oensingen, MP 26) characterized by its slightly larger size, its very brachyodont teeth with long mesolophs/mesolophids and well-developed anterolophs/anterolophids. Also a noticeable peculiarity of the E. molassicus mandible, besides its high ramus horizontalis, is the very large and a little furrowed enamel band on the labial side of its incisor ( Engesser 1987: fig. 5d and cast). In our opinion, based on what we know now on the potential morphological variability of species for the genus Eomys , E. nov. sp. 2 could be either related to E. antiquus or to E. molassicus .
It is worth noting that in the MP 26 level, two clearly different lineages are present: a very brachyodont one, E. molassicus and another with incipient mesodonty, Eomys zitteli known in Mas-de-Pauffié. Their relationships with E. antiquus are not clear at the time.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.