
Vespertiliavus aenigma Ravel, 2016
|
publication ID |
https://doi.org/10.5252/g2016n3a3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:FC07ACBE-03F7-414A-BB64-1BB0711766BF |
|
persistent identifier |
https://treatment.plazi.org/id/03E07259-FF9D-FF96-FF2D-FBF3C85BFDFE |
|
treatment provided by |
Felipe |
|
scientific name |
Vespertiliavus aenigma Ravel |
| status |
sp. nov. |
? Vespertiliavus aenigma Ravel , n. sp.
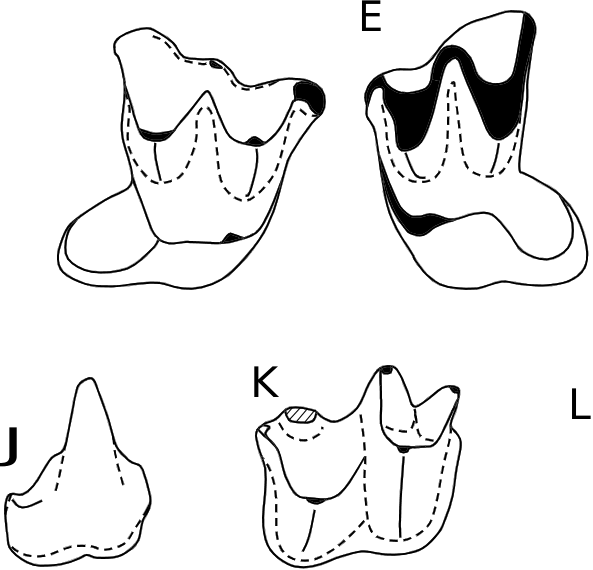
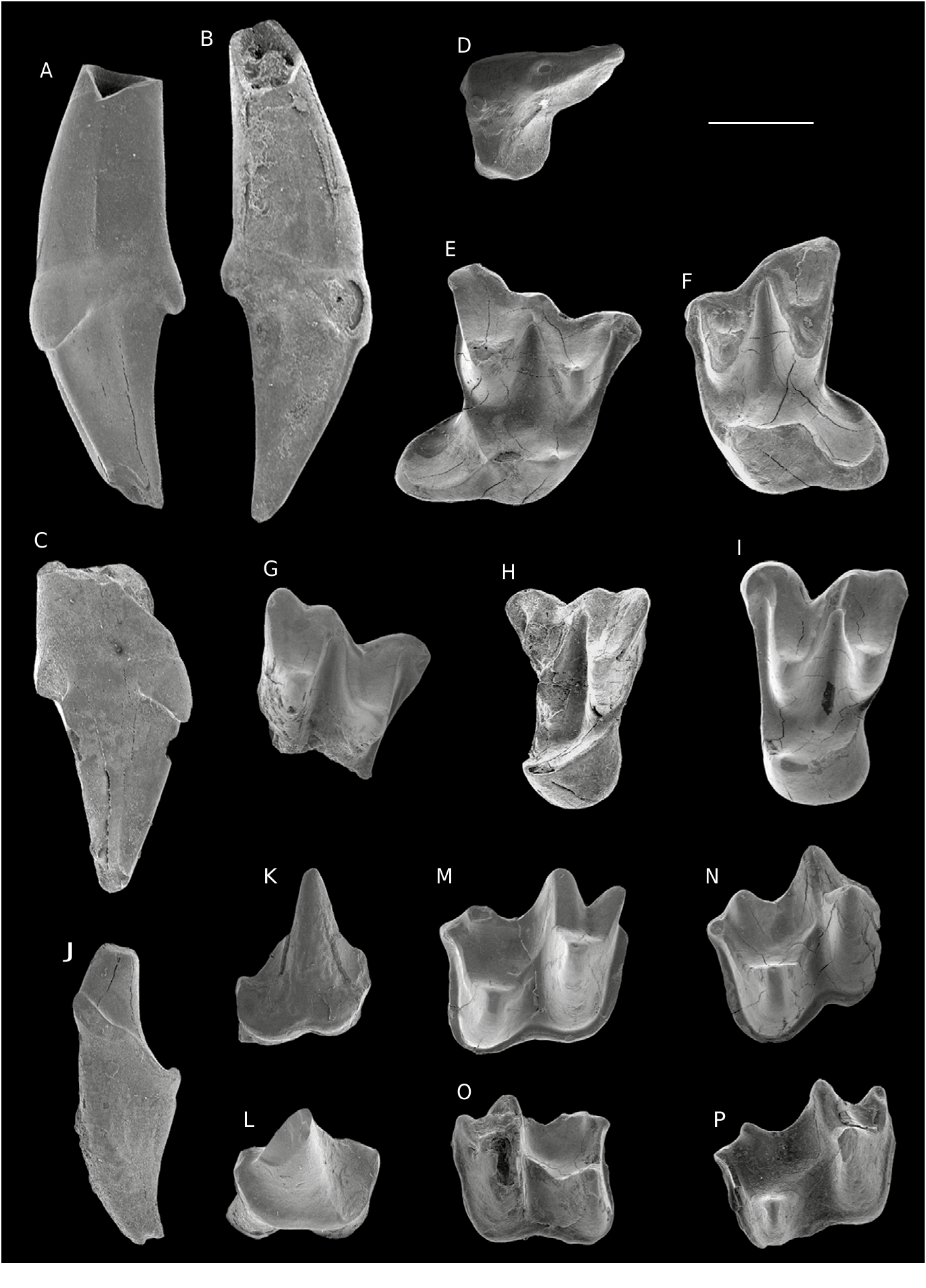
( Figs 14 View FIG , 15 View FIG ; Tableau 5)
HOLOTYPE. — CBI-2-169, M1 droite, Figs 14D View FIG ; 15 E. View FIG
MATÉRIEL EXAMINÉ. — CBI-2-175 (C1 droite; Figs 14A View FIG ; 15 A View FIG ); CBI-2-176 ( Figs 14B View FIG ; 15 B View FIG ) et CBI-2-177 (C1s gauches; Fig.15C View FIG ); CBI-2-174 (P4 gauche; Figs 14C View FIG ; 15 D View FIG ); CBI-2-170 (M1 gauche; Figs 14E View FIG ; 15F View FIG ); CBI-2-171 (ectolophe M2 droite; Figs 14F View FIG ; 15 G View FIG ); CBI-2-173 (M3 gauche); CBI-3-013 ( Figs 14G View FIG ; 15 I View FIG ) et CBI-2-172 (M3s? gauches; Fig.15H View FIG ); CBI-2-205 (c1 droite; Figs 14H View FIG ; 15 J View FIG ); CBI-2-183 (p4 droite; Figs 14J View FIG ; 15 K View FIG ); CBI-2-182 (p4 gauche; Figs 14I View FIG ; 15 L View FIG ); CBI-2-178 ( Figs 14L View FIG ; 15 P View FIG ), CBI-2-179 ( Fig.15N View FIG ), CBI-2-181 ( Figs 14K View FIG ; 15 M View FIG ) et CBI-1-262 (m1/2s droites); CBI- 2-180 (m1/2 gauche; Fig. 15O View FIG ).
ÉTYMOLOGIE. — Du latin « aenigma » qui signifie énigme, en raison de l’originalité des morphologies caractérisant ces spécimens et rendant ainsi l’attribution systématique difficile.
LOCALITÉ TYPE ET ÂGE. — Chambi (CBI) loci 1, 2 et 3, Éocène inférieur terminal-Éocène moyen basal, Djebel Chambi, situé dans la région de Kasserine, Tunisie.
DIAGNOSE. — Vespertiliavus caractérisé par une canine supérieure avec un cingulum très épais et continu, une P4 très étirée longitudinalement, et une M3 possédant un ectolophe qui conserve la postmétacrête et le métastyle semblable au M1/2. Espèce plus grande (environ une fois et demi) que Vespertiliavus kasserinensis Ravel , n. sp.
DESCRIPTION
Le matériel attribuable à cette plus grande espèce du genre Vespertilavus est très fragmentaire. Les canines supérieures ne possèdent qu’un seul tubercule de hauteur modérée et légèrement recourbé vers la partie distale ( Figs 14A, B View FIG ; 15 View FIG A-C). Le cône est constitué d’une face buccale convexe et d’une face linguale légèrement concave. Les deux faces sont délimitées par une crête mésiale et une crête distale plus affutée. Le cingulum, très épais et continu, porte deux bourrelets à la base linguale de la couronne.
La P4 est très étirée longitudinalement mais comprimée buccolingualement ( Figs 14C View FIG ; 15 D View FIG ). La couronne possède un contour triangulaire en vue occlusale dont le côté linguodistal est fortement incurvé vers l’intérieur. La dent est munie d’un unique paracône positionné centrobuccalement. Une crête saillante longiligne (i.e., postparacrête) s’étend depuis le sommet du paracône pour rejoindre le bord buccodistal de la couronne. Cette crête prend une orientation à prédominance distale, et s’incurve très légèrement vers le bord buccal. Le bassin lingual est comprimé transversalement et s’étale sur la moitié mésiolinguale de la couronne. Un fin cingulum lingual cerne le bord lingual du bassin.
La couronne de la M1 CBI-2-169 possède un talon très projeté distalement ( Figs 14D View FIG ; 15E View FIG ). Le bord buccal est incliné mésiolingualement et présente deux ectoflexus de part et d’autre de la région mésostylaire. Le parastyle est légèrement plus lingual que le mésostyle, et nettement plus par rapport au métastyle. Le parastyle est modérément recourbé vers le côté mésial pour former un crochet antérieur. Le métastyle est simple et apparaît buccalement en continuité avec la postmétacrête. Le cingulum buccal, discontinu, est toutefois très épais au niveau du mésostyle. Le mésostyle, situé à la jonction entre la postparacrête et la prémétacrête, est légèrement déplacé lingualement par rapport au bord buccal. Les crêtes de l’ectolophe sont assez divergentes et disposées en «W» ouvert mais légèrement compressé buccolingualement. Le paracône est peu réduit et plus lingual que le métacône. Ces deux tubercules sont séparés par une large et longue protofosse qui est délimitée buccalement par le mésostyle. Le protocône est buccolingualement compressé. Il occupe une position très mésiolinguale tout en restant un peu moins mésial que le paracône. Le protocône est plus comprimé buccolingualement que le métacône et le paracône. La préprotocrête remonte buccalement depuis le sommet du protocône pour rejoindre le précingulum, clôturant ainsi la protofosse mésialement. La postprotocrête est deux fois plus longue que la préprotocrête, et beaucoup plus rectiligne, avec une direction distolinguale depuis le sommet du protocône. Cette crête se termine distalement au niveau d’une triple jonction réalisée avec le cingulum lingual et une crête tranchante qui s’étend sur presque toute la largeur buccolinguale de la couronne depuis le postcingulum. Cette dernière crête marque la limite entre la protofosse et le bassin du talon. Ce dernier est circulaire et se déploie distalement, formant ainsi une structure fortement dissociée du reste de la couronne mais qui participe à une large et longue surface d’occlusion. Son extrémité distale est circulaire et ceinturée par un cingulum lingual modérément épais.
CBI-2-170 ( Figs 14E View FIG ; 15F View FIG ) est interprétée ici comme une M1 de par sa taille, laquelle est similaire à celle de CBI-2-169. La forte inclinaison du bord buccal, la position linguale du parastyle, la projection distale du talon sont des caractères comparables à l’holotype (CBI-2-169, M1; Figs 14D View FIG ; 15 E View FIG ). Cependant ces deux dents diffèrent dans l’agencement et la morphologie de certaines structures. Sur CBI-2-170, la bordure buccale est plus courte et présente un seul ectoflexus, lequel est moins prononcé que sur CBI-2-169. Sur CBI-2-170, la région parastylaire est brisée, ce qui ne permet pas d’évaluer sa morphologie ni sa projection. Distobuccalement, l’ectolophe est plus resserré mésiodistalement, générant ainsi une parafosse et une métafosse plus étendues buccolingualement mais plus compressées mésiodistalement. La postprotocrête est plus courte. Elle rejoint le cingulum lingual, laissant ainsi une protofosse ouverte distalement, qui communique ainsi avec le bassin du talon. Le cingulum lingual apparaît également plus épais. Ces différences peuvent s’expliquer soit par une variation intraspécifique – courante au sein des Vespertiliavus ( Maitre 2014) – soit par la présence ici d’une espèce différente.
Un fragment d’ectolophe d’une M2 ( Fig. 14F View FIG ; 15G View FIG ) possède une taille et une morphologie qui pourrait correspondre à? V. aenigma Ravel , n. sp. Ce dernier possède un bord buccal très légèrement incliné mésiolingualement qui indique une M2 plutôt qu’une M1. Il présente un très léger ectoflexus au dessus du métacône et un autre nettement plus prononcé qui surplombe le paracône. Le parastyle est modérément recourbé et projeté vers bord mésial.
Les M3s? ( Figs 14G View FIG ; 15 H, I View FIG ) ont une largeur équivalente à celle des M1s. L’ectolophe est complet avec un fort ectoflexus au niveau du paracône. À la différence du métastyle qui apparaît comme un simple et petit style, le parastyle forme un crochet buccomésial. Le paracône est plus pincé et plus lingual que le métacône. Le métacône et légèrement réduit. Les post- et préprotocrêtes rejoignent respectivement les pré- et postcinginguli, délimitant ainsi mésialement et distalement la protofosse. Le protocône est réduit et présente un flanc lingual arrondi dépourvu de cingulum. La morphologie de cette dent est peu commune chez les chiroptères. Cependant l’absence de cingulum lingual ne peut correspondre avec une M1 ou une M2 (condition généralisée chez les chiroptères). Le type M3 semble le plus probable. En terme de taille, le matériel référé à? Vespertiliavus aenigma Ravel , n. sp. semble le plus adéquat. Le fort développement de la M3 chez les Emballonuridae en général aurait pu être accentué chez ce taxon. En absence de matériel plus fourni, une telle association reste cependant en suspend.
La canine inférieure est allongée et relativement étroite. La partie la plus apicale de la racine possède une longueur équivalente à la couronne ( Figs14H View FIG ; 15J View FIG ). Cette dernière est constituée d’un unique tubercule (cassé au quart de sa hauteur sur CBI-2-205) présentant une section ovale. Le cingulide n’est visible que sur la face linguale, il est absent au niveau de la base buccale du conide. Le cingulum est fortement incliné distalement. Il effectue un ressaut au niveau de la bordure distale, faisant naître un petit relief qui délimite le bassin de la canine.
La couronne de la p4 présente un contour rectangulaire étroit mais allongé en vue occlusale ( Figs 14I, J View FIG ; 15K, L View FIG ). Sa longueur équivaut à la moitié de celle de m1-2. Elle possède un unique protoconide, très haut et pointu, qui domine la partie mésiale. Une courte crête débute au quart de la hauteur de cette cuspide et s’oriente mésiolingualement, formant ainsi un relief prononcé à l’avant de la couronne. La partie distale est composée d’un bassin modérément allongé mais occupant toute la largeur de la couronne. Une crête distale apparaît depuis le quart de la hauteur du protoconide et rejoint le bord distal délimitant la bordure linguale du bassin du talonide. Un cingulide fin et continu parcourt tout le contour de la dent.
Le trigonide des molaires inférieures est plus court et nettement plus étroit que le talonide ( Figs 14K, L View FIG ; 15M, N, O, P View FIG ). Le paraconide, plus petit que le métaconide, est proclive. Le protoconide, légèrement compressé mésiodistalement, constitue la cuspide la plus volumineuse de la dent. La crête oblique débute depuis la base de la muraille distale du trigonide, au niveau de l’échancrure entre le métaconide et le protoconide. Elle rejoint l’hypoconide (beaucoup plus buccal que le protoconide) en effectuant une forte inflexion buccale. L’hypoconide est faiblement pincé mésiodistalement à son extrémité. La postcristide se projette distolingualement depuis le sommet de l’hypoconide jusqu’à l’hypoconulide, définissant ainsi une structure nyctalodonte. L’hypoconulide est très distal et se situe légèrement plus lingualement que l’entoconide, ce dernier étant élevé et saillant. L’entocristide forme une muraille linguale au bassin du talonide en débutant depuis le sommet de l’entoconide et rejoignant le tiers de la hauteur distale du métaconide. Un fin cingulide est ininterrompu le long des bordures mésiale, buccale et distale de la couronne.
COMPARAISON ET DISCUSSION
Les spécimens décrits ci-dessus présentent une grande variation de forme qui pourrait éventuellement refléter ici la présence de plusieurs espèces.Toutefois, le matériel présente un ensemble de caractères dentaires assez commun chez les Emballonuridae , et plus particulièrement chez le genre fossile Vespertiliavus . Ces caractères sont:
– le cingulum des canines supérieures bien individualisé;
– la compression buccolinguale des P4 et leur contour occlusal triangulaire;
– le parastyle des M1 déplacé lingualement, entraînant une inclinaison modérée à forte du bord buccal;
– le bassin du talon très étendu distalement et en largeur; – la postprotocrête orientée distalement rejoignant parfois l’hypocône ou le cingulum lingual;
– la connexion entre la protofosse et le bassin du talon, conférant à la première et la seconde molaire une large surface d’occlusion (pour CBI-2-170; Figs 14E View FIG ; 15F View FIG );
– p4 à contour rectangulaire étroit mais allongé;
– le protoconide de p4 très haut et pointu;
– la compression mesiodistale du trigonide formant un «V » refermé en vue occlusale;
– l’aspect gracile de l’entoconide, lequel est pointu et incliné distalement;
– l’extension du talonide plus large que le trigonide et aussi long;
– la M3 bien développée, munie d’un métacône bien défini. La morphologie et l’orientation de la postprotocrête ainsi que l’isolement du bassin du talon (observées sur CBI-2-169, l’holotype; Fig. 14 D View FIG ; 15E View FIG ), représente un agencement peu commun au sein des Emballonuridae . Parmi le matériel de comparaison, seul Saccolaimus peli Temminck, 1853 présente un bassin isolé (uniquement sur la M1), mais cet isolement est réalisé par l’extension du métalophe qui rejoint la postprotocrête, et n’implique donc pas les mêmes structures. Dans l’abondant matériel du Quercy attribuable au genre Vespertiliavus (collection hébergé à l’ISE-M, matériel provenant des niveaux repères MP13 à MP26), certains spécimens présentent une courte postprotocrête orientée distobuccalement, entraînant une rupture avec le cingulum lingual. Cette variation, que l’on retrouve au sein d’une espèce ( V. bourguignati Filhol, 1877 ; La Bouffie MP17a), n’aboutit cependant jamais à une ferme - ture franche de la protofosse (la postprotocrête s’interrompt avant de rejoindre la base du métacône). Sur certains aspects, notamment au niveau de l’ectolophe, CBI-2-169 se rapproche des Mixopterygidae et plus précisément du genre Mixopteryx Maitre, Sigé & Escarguel, 2008 ( Maitre et al. 2008). La particularité des Mixopterygidae est de combiner des caractères emballonuroïdes avec des caractères rhinolophoïdes. Ces caractéristiques n’ont pas permis de rapprocher cette famille de l’un des grands ensembles de chiroptères actuels. CBI-2-169 possède un double ectoflexus, un mesostyle bien projeté buccalement, et un parastyle déjeté mésialement. Cette suite de caractères s’observe également chez Mixopteryx . Cependant, les molaires inférieures de? Vespertiliavus aenigma Ravel , n. sp. diffèrent radicalement des rhinolophoïdes et présente un patron dentaire typique des Emballonuridae . En effet, chez les emballonuroïdes, tout comme chez? V. aenigma Ravel , n. sp., le trigonide est fermé, l’entoconide est haut et penché distalement, et le talonide est plus large que le trigonide. En revanche, chez les rhinolophoïdes, et notamment chez les Mixopterygidae , les molaires inférieures ont un trigonide ouvert, l’entoconide est peu élevé et arrondi, le talonide est réduit, et les cuspides sont larges.
L’autre fait remarquable relatif à? V. aenigma Ravel , n. sp. est la morphologie de ses M3s. Elles ont une bordure linguale arrondie et dépourvue de cingulum. Cependant la morphologie et la taille de l’ectolophe de ces dents sont similaires aux conditions observées sur les M1-2s. Ces similitudes incluent des critères morphologiques attribuables à un Emballonuridae : ectoflexus profond au niveau du paracône, resserrement des crêtes de l’ectolophe avec un développement transverse de ce dernier, cingulum buccal bien développé, protofosse étendue, et protocône déjeté vers l’avant. Même si les M3s de la majorité des espèces d’Emballonuridae actuels (par ex.: Peropteryx macrotis Wagner, 1843 , Rhynchiscus naso Maximilian, 1820 , Rhynchonycteris naso Maximilian, 1820 , Saccopteryx leptura Schreber, 1774 et S. bilineata Temminck, 1838 , Emballonura atrata Peters, 1874 , Diclidurus albus Wied-Neuwied, 1820 , et Cormura brevirostris Wagner, 1843 ) ne sont pas aussi complexes, il existe des cas où le métacône peut-être bien individualisé, avec une largeur de dent similaire à celle des M2s. Ces caractéristiques sont également observables chez les formes fossiles Tachypteron franzeni et l’ensemble des Vespertiliavus . Il est donc possible que ce morphotype caractérise l’espèce? Vespertiliavus aenigma Ravel , n. sp.
B DG H E F
C J
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.