
Xylopertha praeusta ( Germar, 1817 )
|
publication ID |
https://doi.org/10.5852/ejt.2017.380 |
|
publication LSID |
lsid:zoobank.org:pub:F80DE451-78CA-406A-A879-E7BD08279ECD |
|
DOI |
https://doi.org/10.5281/zenodo.5695411 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF879F-9F22-A03E-1836-FC66FBF3FD25 |
|
treatment provided by |
Plazi |
|
scientific name |
Xylopertha praeusta ( Germar, 1817 ) |
| status |
|
Xylopertha praeusta ( Germar, 1817) View in CoL
Figs 1E–F View Fig. 1 , 2B View Fig. 2 , 3B View Fig. 3 , 4C View Fig. 4 , 5C–D View Fig. 5
Apate praeusta Germar, 1817: 226 .
Xylopertha appendiculata Lucas, 1849: 466 . Synonymy: Jacquelin du Val 1860: 164.
X ylopertha dunensis Rai & Chatterjee, 1964: 122 View in CoL . Synonymy: Borowski & Singh 2017: 198.
Xylonites praeustus – Lesne 1901: 581.
Xylopertha praeusta View in CoL – Rai & Chatterjee 1964: 124.
The type material of X. praeusta was not located, but the species is well known in the Mediterranean area. The senior author has examined numerous specimens determined by Lesne and Vrydagh, and numerous other specimens in IRSNB, LYL, MAIC, MNB, NHMUK, SDEI, SNSD and ZMHB, from various countries in southern Europe and North Africa. Borowski & Singh (2017) examined the type series of Xylopertha dunensis deposited in IFRI, and synonymised X. dunensis with X. praeusta . They state that the specimens in the type series came in fact from Tulon [ sic] (correctly Toulon), S-France. It has become apparent that K. Rai falsified the type location of several species that he described ( Borowski & Singh 2017). India should, therefore, be excluded from the distribution of X. praeusta . If the species does occur in India, it is almost certainly introduced.
Diagnosis
The species is distinguished from X. elegans sp. nov. and X. retusa by the presence of a spine on each elytron in both sexes ( Fig. 1 View Fig. 1 ). It is distinguished from X. reflexicauda by the position of the spine, which is on the dorso-lateral margin of the declivity in X. praeusta , but on the middle of the upper margin of the declivity in X. reflexicauda . Differences in the abdomen and genitalia are illustrated in Figs 4 View Fig. 4 and 5 View Fig. 5 .
Material examined
ITALY: 1 ♂, [Corsikac. Ug., Corte , 28 May 20, 11, F/o Bmann, Tavignano-Tal] ( NME).
FRANCE: 1 ♂, 1 ♀, [ Larale Valley, Pyrenees , J. Kettere leg.] ; 1 ♀, [A. Melie L/B, LE/ 5-6-1960, ex coll. A. Jacobs ( IG: 28.548 )] .
MOROCCO: 1 ♂, [ Col du Zad , 13 Jul. 1959, M. Vazquez leg.] ; 1 ♂, [ Bab-Bou-Idir env., 34°4′ N, 4°7′ W, 28 May–3 Jun. 1999, Tazzeka N.P. , I. Smatana leg.] ( LYL) GoogleMaps .
The specific identity of more than 260 further specimens was checked in the following museums: IRSNB, MAIC, MIZPAN, MNB, MTM, NHMUK, NMBS, NMPC, NMS, NMW, SDEI, SNSD, ZNHB and ZSM, but detailed locality data were not recorded. All specimens came from the distributional area given below.
Description
BODY. 4–8 mm long, about 2.5–2.8 times as long as wide, stout, parallel-sided. Body shiny, head, pronotum, ventral side and femora black; elytral disc dark brown, darker toward the declivity; tibiae dark brown, antennae and tarsi paler.
HEAD. Clypeus finely and densely punctured, convex along its anterior margin, its median truncation as wide as base of labrum. Frons with puncturation less dense than clypeus, covered by fairly long, upwardly directed hairs. First and second segments of antennal club slightly elongate. Fifth antennal segment often nicked on external side, but this is not constant, nor particular to one sex. Pronotum about 1.1 times wider than long, narrowed in anterior third, without long hairs antero-laterally, area above the head finely punctured.
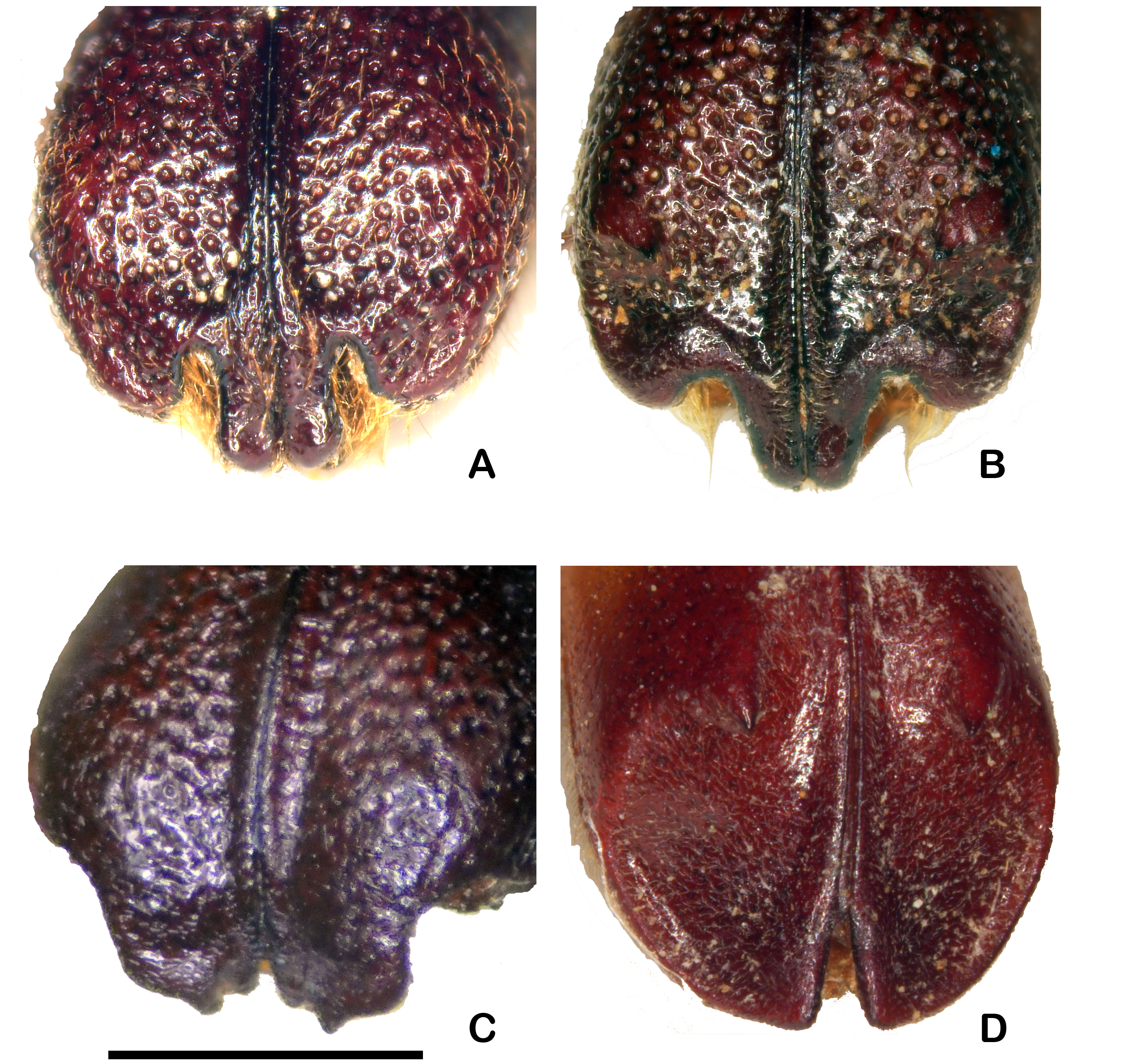
ELYTRA. Puncturation dense and strong on whole of disc; elytral declivity with a tubercle-like spine with a pointed apex located on dorso-lateral margin of declivity of each elytron ( Figs 1E–F View Fig. 1 , 3B View Fig. 3 ); pubescence of apical declivity short, erect, reddish, especially noticeable near upper margin where formed of longer hairs.
LEGS. External face of anterior tibiae narrowly grooved, widened to apex. Second and third segments of anterior tarsi scarcely wider than the following ones.
Male
ELYTRA. Apical declivity larger than in female, covered by an exceedingly fine and dense puncturation, and often wrinkled in its upper part; elytral spine robust, directed inwardly, strongly bulbous on outer side at base, narrow and very pointed at apex, below it a broad flange beginning at sutural angle and extending at sides to about level of lateral spine; sutural angles not projecting, rounded at apex ( Fig. 1E View Fig. 1 ).
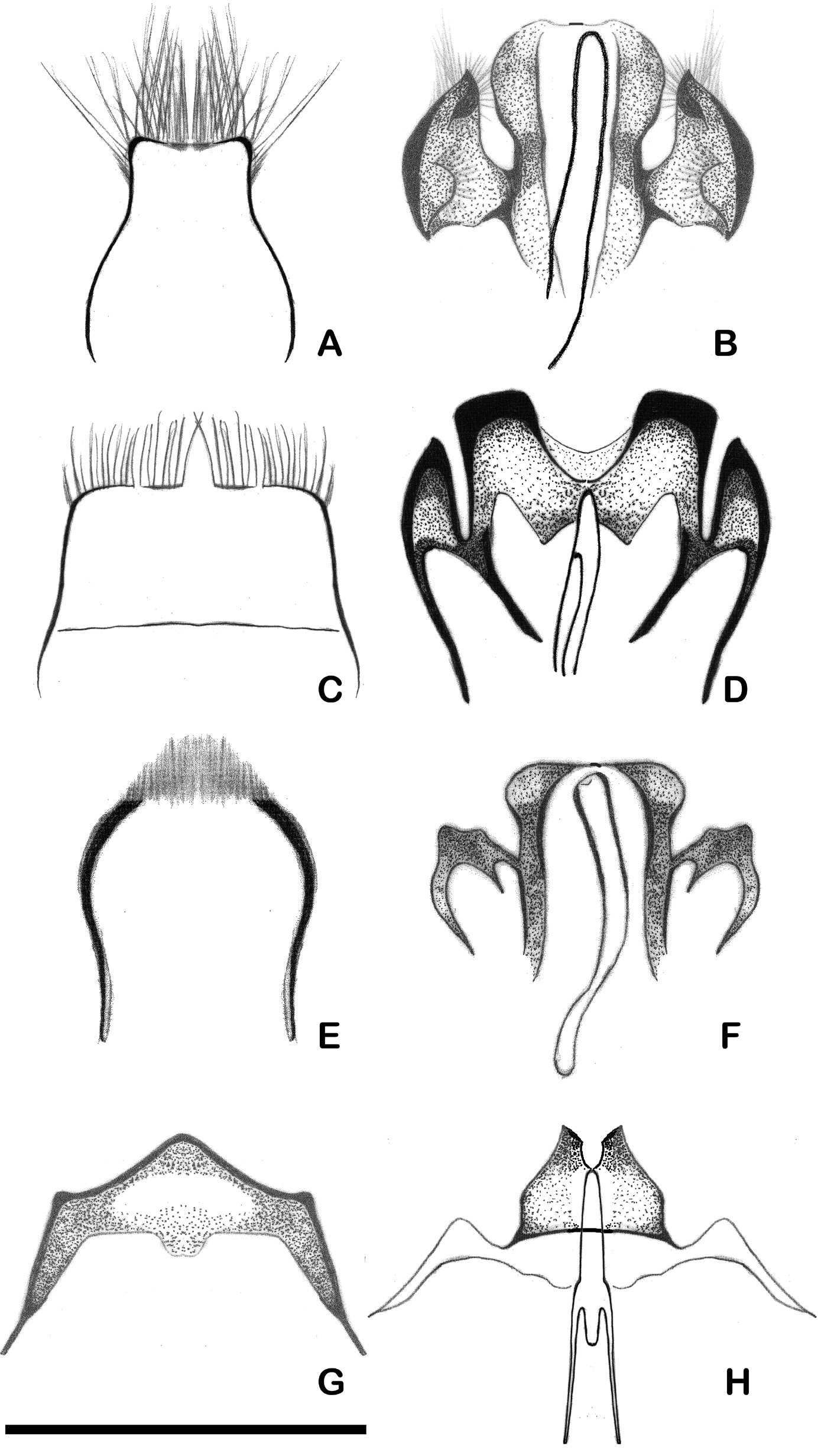
ABDOMEN. Anterior margin of 8th tergite wider and fringed by shorter erect hairs than in X. retusa . Genitalia more strongly sclerotised than in X. retusa , with pair of ventrally-directed processes with pointed tips next to inner lobes which are separated by a U-shaped emargination in middle. Aedeagus simple, without long apophyses ( Fig. 5C–D View Fig. 5 ).
Female
ELYTRA. Declivity less distinctly truncate than in the male, strongly punctured, the lateral spines smaller, or replaced by tubercles; suture inflated on the declivity ( Fig. 1F View Fig. 1 ). Apical emargination of elytra broad, the emargination largely filled by a pair of ventrally-directed, extended processes with rounded tips next to suture ( Fig. 3B View Fig. 3 ).
ABDOMEN. Last visible segment of abdomen densely pubescent on sides, not grooved in middle; posterior margin of segment armed by two dorsally recurved large median teeth ( Fig. 4C View Fig. 4 ).
Distribution
Southern Europe, North Africa, Israel, Turkey.
Biology
The species has been recorded from the following genera and families of trees: Viburnum L. ( Adoxaceae E.Mey. ), Mangifera L., Pistacia (Anacardiaceae) , Acacia Mill. (Fabaceae) , Castanea Mill. , Quercus L. ( Fagaceae ), Ficus L. ( Moraceae ), Eucalyptus L’Hér. ( Myrtaceae Juss. ), Populus L. ( Salicaceae ), Vitex L. ( Vitaceae Juss. ) ( Lesne 1901; Rai & Chatterjee 1964; Akşit et al. 2005; Buse et al. 2013). However, it develops primarily in oaks ( Quercus ) ( Bahillo de la Puebla et al. 2007; Nardi & Mifsud 2015). In Europe, adults emerge in May–July ( Lesne 1901), but are active until November in Israel ( Buse et al. 2013). The species has diurnal habits. The gallery system is typical of the genus. Larvae overwinter and complete their development in the spring of the following year ( Lesne 1901).



| LYL |
LYL |
| NME |
Sammlung des Naturkundemseum Erfurt |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Bostrichidae |
|
Family |
|
|
SubFamily |
Bostrichinae |
|
Tribe |
Xyloperthini |
|
Genus |
Xylopertha praeusta ( Germar, 1817 )
| Liu, Lan-Yu & Beaver, Roger A. 2017 |
dunensis
| Rai & Chatterjee 1964: 122 |
Xylopertha appendiculata
| Lucas 1849: 466 |
Apate praeusta
| Germar 1817: 226 |