
Oedichirus misionesiensis, Herman, 2013
|
publication ID |
https://doi.org/ 10.1206/816.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03DF8794-7D79-D102-FD5F-53B9FDFE0347 |
|
treatment provided by |
Felipe |
|
scientific name |
Oedichirus misionesiensis |
| status |
sp. nov. |
Oedichirus misionesiensis View in CoL , new species
Figures 40 View Figs , 172–179 View Figs View Figs
TYPE MATERIAL: Holotype: Male. ‘‘ARGEN- TINA: Prov. Misiones, 15 km SE Puerto Iguazo[sic], 30.XII.1990 /FMHD # 90-312, forest, S & J Peck, 90-141, treebase litter FIELD MUSEUM/ Oedichirus P.N. Thomas det. 1993/ Holotype Oedichirus misionesiensis Herman. ’’ Deposited in the Field Museum of Natural History, Chicago. (Left metafemur, metatibia, and metatarsus are missing.)
TYPE LOCALITY: Argentina: Misiones: 15 km SE Puerto Iguazú (25 ° 419S, 54 ° 289W, 240 m).
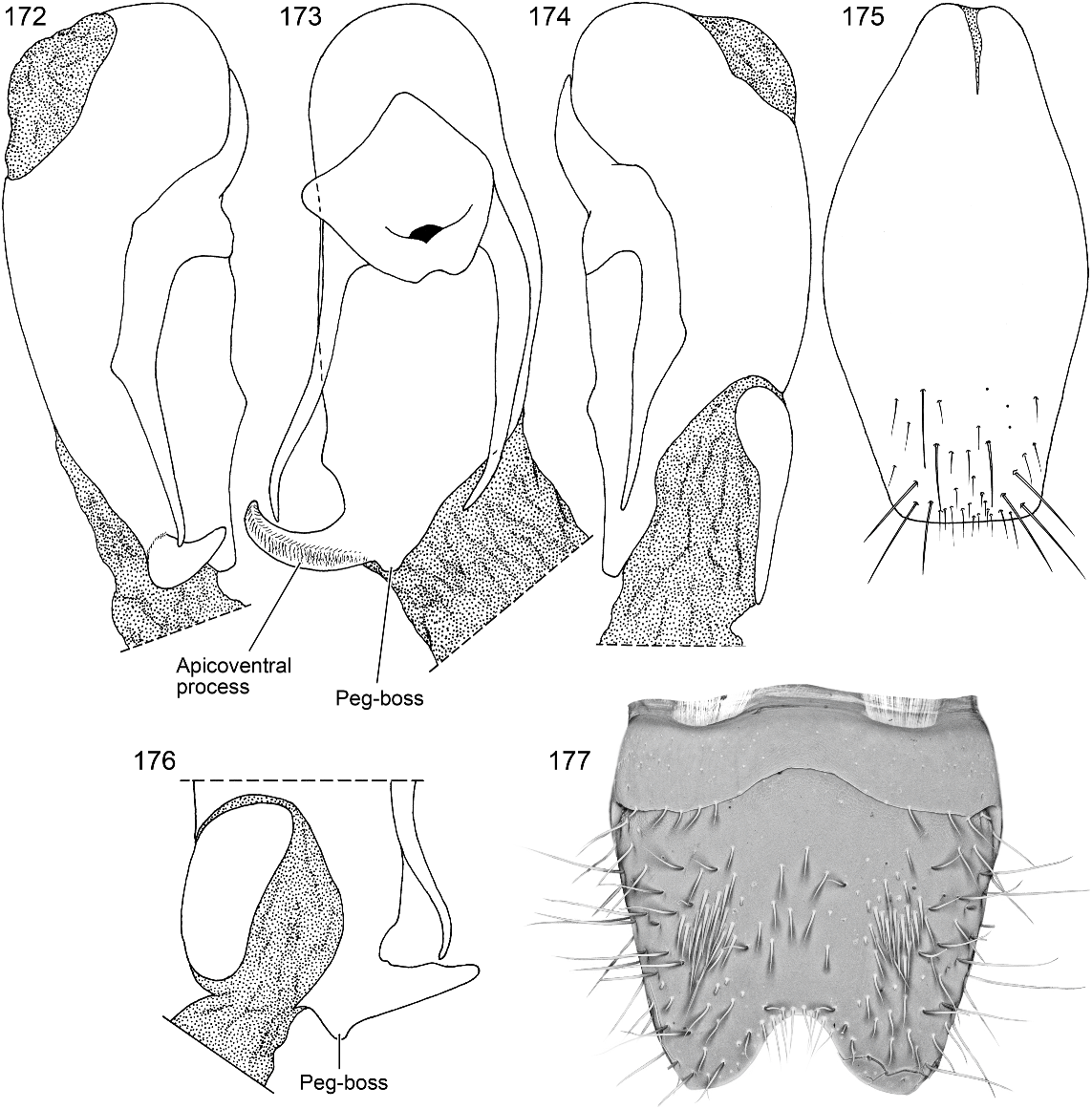
DIAGNOSIS: Oedichirus misionesiensis , part of a group of narrowly separated species that includes O. apiculus , O. glabrihamus , and O. speculifrons , can be separated from them all by the cylindrical peg-boss on the posterior margin of the ventral sclerite near the left base of the aedeagal apicoventral process (fig. 173). Although the size of the basal boss on the apicoventral process of O. speculifrons varies, it differs from that of O. misionesiensis in that it is ventrally directed and manifests as a broadly rounded, undistinguished tumescence. It is separated from O. apiculus by the deeper emargination of sternum VIII (cf. figs. 177 and 46); O. apiculus has a spiniform process on the right dorsal edge of the ventral sclerite near the base of the apicoventral process (figs. 41, 44), O. misionesensis does not (fig. 72). Sternum VIII is similar in O. misionesiensis (fig. 177) and O. speculifrons (figs. 208, 209), but subtle differences of the median depression and associated subapical boss of the latter species will separate them. For O. misionesiensis (fig. 177) the median depression is more shallow and less distinct, the ridge marking the lateral side is feeble, the slope from the lateral side of the depression is slight, the subapical boss is a polished, feeble swelling at the lateroapical corner of the median depression, and the surface between the corners is almost flat. For O. speculifrons (figs. 208, 209) the median depression is deeper, the ridge marking the lateral side is well developed posteriorly, and the subapical boss is higher, more prominent, and the depression between them distinct. As is true for others of this complex of species, the males of O. misionesiensis can be separated from other New World species by the deep, basally rounded, symmetrical emargination of sternum VIII (fig. 177) and the rightwardly directed, stout apicoventral process of the aedeagus (figs. 172, 173).
DESCRIPTION: Length: 7.5 mm. Length of head: 0.7 mm. Width of head: 1.1 mm. Pronotal length: 1.2 mm. Pronotal width: 1.1 mm. Elytral length: 0.8 mm. Elytral width: 1.1 mm.
Body black with scattered dark reddishbrown infusions. Legs yellowish brown, with pale reddish brown coxae; femorotibial joint without dark maculation.
Head about three fifths wider than long (HW/HL 5 1.6). Frontoclypeal ridge incomplete with medial ends widely separated. Dorsal surface without V-shaped depression; punctation coarse, moderately dense, evenly distributed, but absent adjacent to frontoclypeal region and nuchal groove. Labrum quadridentate; surface without tubercle near submedial denticle.
Pronotum about a tenth longer than wide (PL/PW 5 1.1). Pronotum polished with coarse, moderately dense punctation and with moderately large impunctate spots; submedial region with irregular cluster of punctures, but without distinct punctate groove. Elytra about two fifths wider than long (EW/EL 5 1.4); surface slightly convex and coarsely punctate.
Abdominal terga III to VIII coarsely and densely punctate and without transverse rows. Segment III without paratergite; paratergal carina present, extending for about half of length of segment, and moderately well developed beyond spiracle. Tergum III without median point extending from transverse basal ridge. Tergum VIII with truncate posterior margin; transverse basal ridge broadly curved and without median point. Tergum IX with lateroapical process about one fifth longer than midbasal length of tergum (LLaP/L9 5 1.2), slightly bent ventrally, and slightly divergent from other process; ventromedial margin without posteriorly directed spur (cf. fig. 158).
MALE: Sterna VI and VII unmodified. Sternum VIII (fig. 177) with deep, wide, symmetrical emargination of posterior margin; emargination about one eighth of length of segment, wider than deep, and base broadly rounded; subapical surface not translucent and posterior margin not membranous; surface with broad, feeble, nearly flat depression extending from near base of emargination to transverse basal ridge; surface with feeble, polished swelling on lateroapical corner of median depression; median depression without distinct ridge on lateral side extending anteriorly from lateroapical corner, but lateral edge polished and slightly sloped; median depression with moderately dense punctation basally and impunctate apically; posterior end of depression beveled adjacent to emargination and with transverse row of closely spaced setae; surface laterad of median depression with shallow depression and with dense cluster of slender, spinelike setae just proximad of depression; surface distad of lateral depression with curved row of setate punctures; sternal surface without comb; transverse basal ridge with median portion broadly and strongly curved anteriorly and without median point. Tergum IX with moderately long, moderately wide process on anterior margin of anteroventral angle. Sternum IX (fig. 175) slightly asymmetrical; anterior margin moderately wide, slightly emarginate, and with median membranous notch; posterior margin nearly truncate; lateral margin broadly rounded.
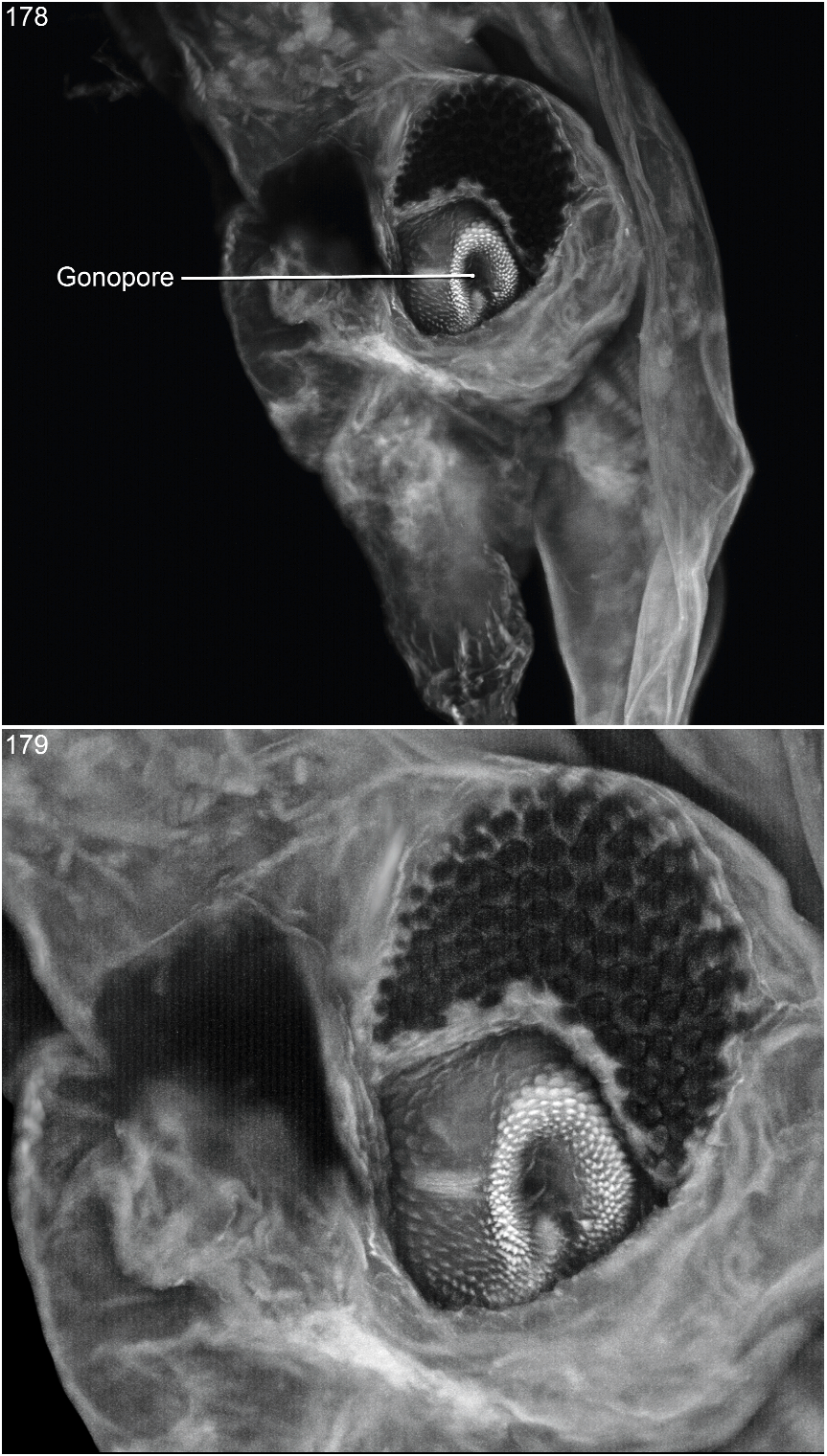
Aedeagus asymmetrical (figs. 172–174). Ventral sclerite with apicoventral process extending laterally from right side of apical margin (fig. 173); apicoventral process moderately long, anteroposteriorly flattened, thick basally and strongly tapered to lateroanteriorly curved, acute apex; ventral sclerite with large, tapered, cylindrical, apically truncate peg-boss at base of left side of apicoventral process (figs. 173, 176), with small tumescence on right ventral side proximad of base of apicoventral process, and without spiniform process or pointed lobe on right dorsal margin dorsad of base of apicoventral process (fig. 172). Parameres long, tapered, moderately broad basally and slender apically and reaching to near posterior margin of ventral sclerite; base fused to median lobe, apical portion free of median lobe. Internal sac (figs. 178, 179) largely membranous; apical region with gonopore surrounded by short, apically acute and apically rounded cuticular processes (fig. 178); ventral side with dense cluster of short, apically round pegs lateroposteriad of and to right of gonopore (fig. 179); dorsal surface with short, robust, spinelike spur subapically.
FEMALE: Unknown.
ETYMOLOGY: This, the first Argentine species of the genus, is named for Misiones province, from which it was collected.
DISTRIBUTION: Known only from the northeastern most province of Argentina, Misiones (fig. 40).
REMARKS: Oedichirus misionesiensis is the first species of the genus reported for Argentina. It is similar and related to O. apiculus , O. glabrihamus , and O. speculifrons , all found in southeastern Brazil at Nova Teutonia, about 350 km (by air) east of Puerto Iguazú, Argentina, which is near the type locality of O. misionesiensis . The Argentine species was collected in leaf litter at the base of a tree. Habitats for the three Brazilian species have not been reported.
The internal sac of O. misionesiensis has three heavily sclerotized structures, none of which are well illustrated in the confocal images (fig. 178, 179). For the specimen of this species the structures simply did not fluoresce as well as homologous structures of other species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.