
Anagrus (Anagrus) epos Girault
|
publication ID |
https://doi.org/ 10.5281/zenodo.194665 |
|
DOI |
https://doi.org/10.5281/zenodo.6209437 |
|
persistent identifier |
https://treatment.plazi.org/id/03DE879A-815E-FF87-C9B6-32C9FB42FE9F |
|
treatment provided by |
Plazi |
|
scientific name |
Anagrus (Anagrus) epos Girault |
| status |
|
Anagrus (Anagrus) epos Girault View in CoL
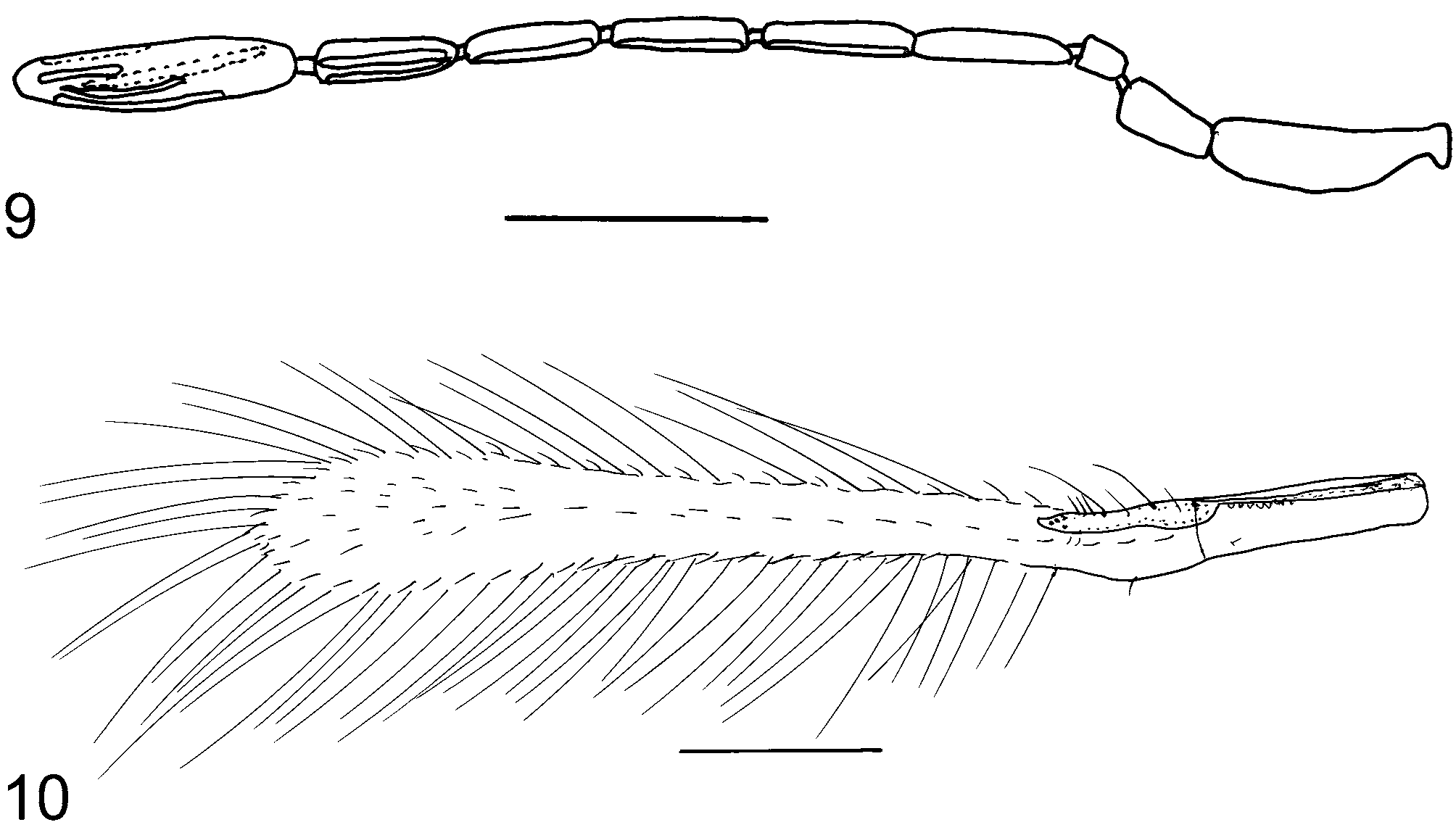
( Figs 9, 10 View FIGURES 9, 10 )
Anagrus epos Girault 1911: 292 View in CoL –293; Triapitsyn 2006: 7 View Cited Treatment –9; Morse & Stouthamer 2007: 94 –96. Anagrus (Anagrus) epos Girault View in CoL : Triapitsyn 1998: 100–100 (in part, type material and non-type specimens from Illinois only).
Type locality. Centralia, Marion Co., Illinois, USA.
Material examined. The specimens listed by Triapitsyn (1998) from Illinois only and Triapitsyn (2006) from Minnesota and also the following specimens: USA, California, Riverside Co., Riverside, University of California at Riverside quarantine laboratory, from colony on Homalodisca vitripennis (Germar) eggs on leaves of Euonymus japonica , reared during vi–ix.2004 by V.V. Berezovskiy & S.V. Triapitsyn and from ix.2004 to xii.2006 by R. Krugner; originally from: USA, Minnesota, Clay Co., ca. 4 mi. SEE of Glyndon, Bluestem Prairie (Nature Conservancy Preserve, moist tallgrass prairie), 46.85521°N 96.47353°W, 31.v– 1.vi.2004, R.A. Rakitov (ex. egg masses of Cuerna fenestella Hamilton on Solidago sp. and Zigadenus sp.; emerged in UCR quarantine 8–14.vi.2004, collected and colony established by S.V. Triapitsyn & V.V. Berezovskiy), numerous females and males in ethanol [ UCRC].
Female diagnosis. F3 of antenna ( Fig. 9 View FIGURES 9, 10 ) usually with a mps; basal one-third of forewing disc beyond venation with one well-defined, complete longitudinal row of setae in both sexes ( Fig. 10 View FIGURES 9, 10 ); forewing 7.9–8.6x as long as wide; ovipositor 2.8–3.1x as long as protibia length; external plate of ovipositor with 3 distal setae.
Hosts. Cuerna fenestella Hamilton the natural host in Minnesota ( Triapitsyn & Rakitov 2005; Triapitsyn 2006), Homalodisca vitripennis (Germar) ( Triapitsyn 2006) , and several other factitious hosts listed by Krugner et al. (2007).
Comments. The record of A. epos from New Mexico (Triapitsyn 1998) was mostly likely erroneous due to a misidentification of a very similar species whose identity is not clear, although it may belong to either A. vulneratus or A. sp. near vulneratus . Records of A. epos from Baja California and Sonora are referred to below as A. sp. near vulneratus . The identities of the specimens of A. epos reared from eggs of Erythroneura aclys McAtee and E. bistrata McAtee in Kentucky and of those reared from eggs of E. bistrata and E. comes (Say) in New York (Triapitsyn 1998) need to be verified using molecular methods.
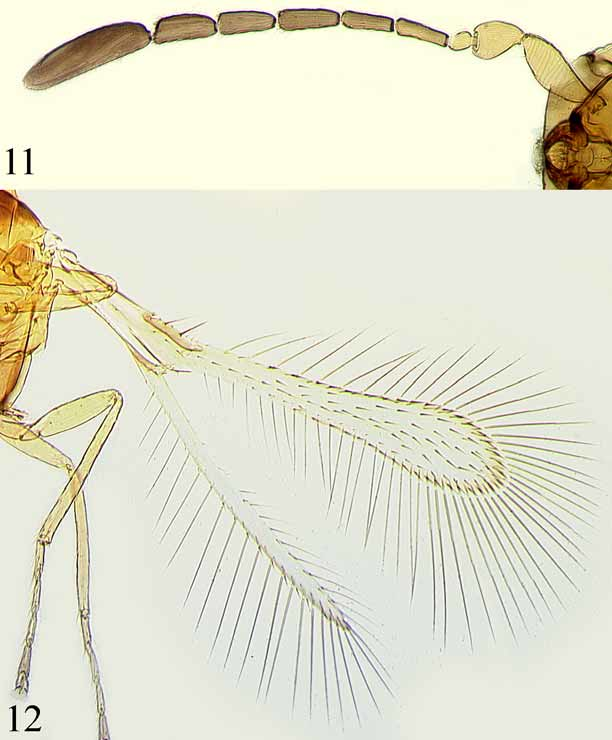
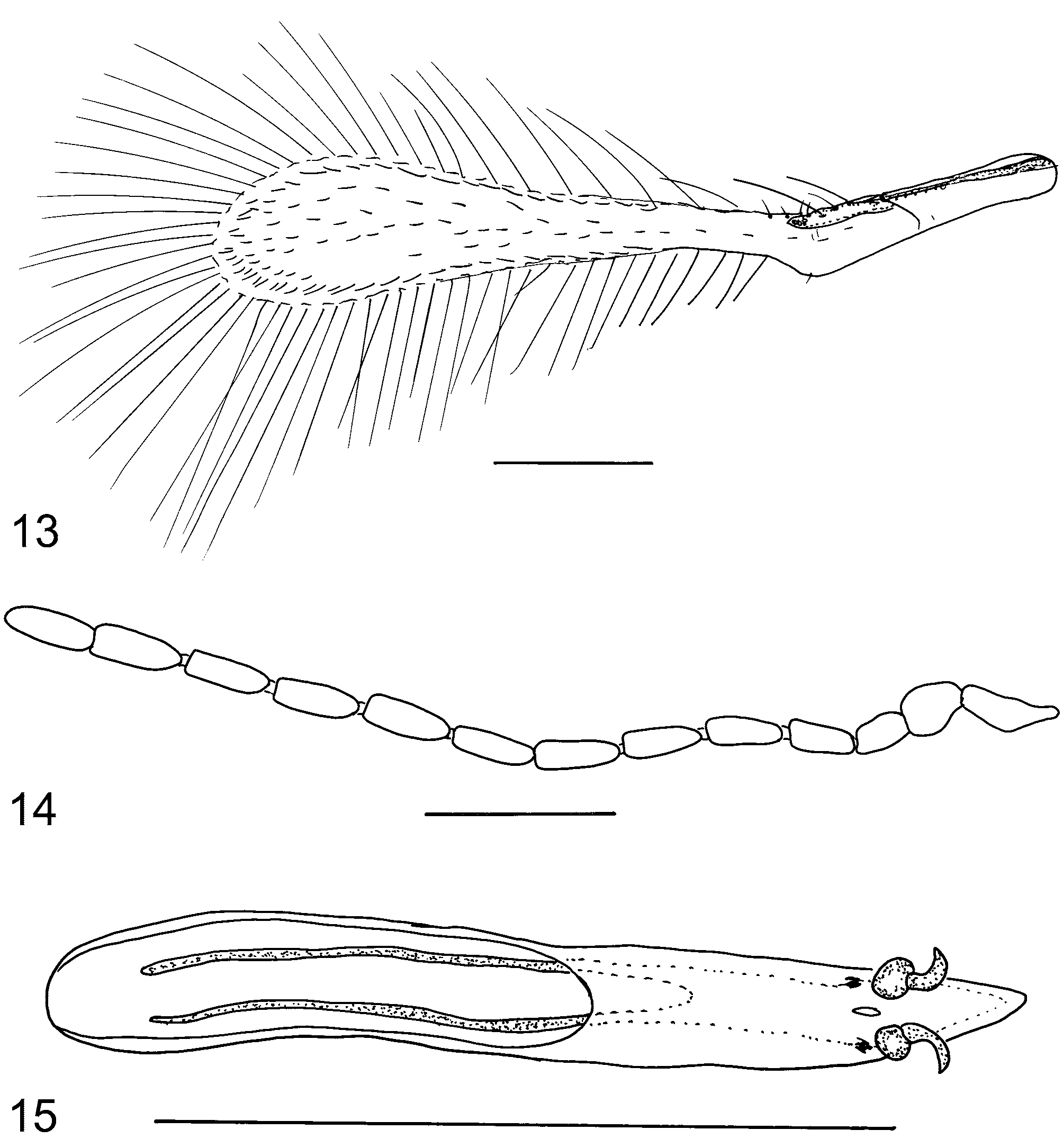
Anagrus (Anagrus) vulneratus Triapitsyn , sp. n. ( Figs 11–15 View FIGURES 11, 12 View FIGURES 13 – 15 )
Anagrus epos Girault View in CoL : González et al. 1988: 23 –25 (misidentification, in part [specimens from Grand Junction, Colorado]).
Anagrus (Anagrus) epos Girault : Triapitsyn 1998: 100–103 (misidentification, in part [specimens from Grand Junction, Colorado]).
Anagrus View in CoL new species: Morse & Stouthamer 2007: 95.
Type material. Holotype female [ UCRC ENT 014814] on slide labeled: “ USA: Colorado, Mesa Co., Grand Junction, Colorado State University Western Colorado Research Center – Orchard Mesa, 3168 B1/2 Road [39°02’31’’N 108°27’58’’W, 1450 m], 29.viii.2006, S.V. Triapitsyn. Emerged 3.ix.2006 at University of California, Riverside quarantine from wine grape leaves infested with Erythroneura vulnerata Fitch S &R # 06–28–01 ”. Paratypes: same data as holotype, 1 female and 1 male on slides [ UCRC]; same data except emerged 30.viii.2006, 1 female on point [ UCRC]; same data except emerged 1.ix.2006, 1 male on point [ UCRC]; same data except emerged 4.ix.2006, 2 females on points [ CNCI, USNM]; same data except emerged 5.ix.2006, 7 females and 2 males on points [ UCRC]; same data except emerged 7.ix.2006, 3 females on points and 1 male on slide [ UCRC]. USA, Colorado, Mesa Co., Grand Junction: 16.vii.1986, D. González (from eggs of Erasmoneura vulnerata (Fitch) on grape leaves), 10 females on slides [ UCRC]; Colorado State University Western Colorado Research Center – Orchard Mesa, 3168 B1/2 Road, 39°02’31’’N 108°27’58’’W, 1450 m: 29.viii.2006, S.V. Triapitsyn (sweeping wine grapes [nursery] infested with E. vulnerata ), 1 male on point [ UCRC]; 4.ix.2007, S.V. Triapitsyn (emerged 14.ix.2007 at University of California, Riverside quarantine from wine grape [nursery] leaves infested with E. vulnerata , S&R # 07–39–01), 1 female on slide [ UCRC] (molecular voucher PR–08–192).
Additional material examined (all in ethanol, UCRC). Same data as holotype except emerged 2.ix.2006, 2 females and 1 male; emerged 5.ix.2006, 1 female; emerged 6.ix.2006, 2 females. USA, Colorado, Mesa Co.: Grand Junction, Colorado State University Western Colorado Research Center – Orchard Mesa, 3168 B1/2 Road, 39°02’31’’N 108°27’58’’W, 1450 m, 4.ix.2007, S.V. Triapitsyn (emerged 7–12.ix.2007 at University of California, Riverside quarantine from wine grape [nursery] leaves infested with E. vulnerata , S&R # 07–39– 01), numerous females and males. Palisade, Garfield Estates vineyard, 29.viii.2006, S.V. Triapitsyn (emerged 6.ix.2006 at University of California, Riverside quarantine from wine grape leaves infested with E. vulnerata , S & R # 06–28–03), 1 female.
Description. FEMALE. Length 430–590 µm. General body color yellow to light brown except transverse trabecula, stemmaticum, anterior half of mesoscutum, and basal metasomal terga notably darker (brown), eyes and ocelli pink; appendages: scape, pedicel, and F1 light brown, the remaining flagellar segments brown, legs light brown.
Antenna ( Fig. 11 View FIGURES 11, 12 ) with scape 2.6–2.8x as long as wide; F1 subglobular or subcylindrical, less than half length of pedicel; F2 and F3 subequal (F3 usually slightly shorter) and each a little shorter than following funicular segments; F4–F6 subequal (F5 usually slightly shorter than F4 and F6); F1 and F2 without mps, F3 usually without mps but occasionally with 1 mps on one antenna, F4 and F5 with 1, F6 with 2 mps each; clava 3.0–3.3x as long as wide, a little longer than combined length of the two preceding segments, with 5 mps.
Mesosoma about 0.7x as long as metasoma. Mesoscutum with a pair of submedian adnotaular setae. Forewing ( Figs 12 View FIGURES 11, 12 , 13 View FIGURES 13 – 15 ) 6.3–6.7x as long as wide; disc hyaline, with 1 complete row of setae extending from apex of venation to wing apex and 2–4 additional, irregular rows of discal setae, usually leaving a small bare area in the broadest part of disc near posterior margin ( Fig. 13 View FIGURES 13 – 15 ) but often without a distinct bare area ( Fig. 12 View FIGURES 11, 12 ); distal macrochaeta 1.8–1.9x as long as proximal macrochaeta; longest marginal seta 2.2–2.7x greatest wing width. Hind wing ( Fig. 12 View FIGURES 11, 12 ) about 22x as long as wide; longest marginal seta about 6.6x greatest wing width.
Ovipositor anteriorly extending to mesophragma and posteriorly exserted beyond apex of gaster by 0.11– 0.14x own length. External plate of ovipositor with 2 or 3 distal setae. Ovipositor 2.5–2.7x as long as protibia length.
Measurements (µm) of the holotype. Body: 590; ovipositor: 279. Antenna: scape: 70; pedicel: 34; F1: 14; F2: 40; F3: 39; F4: 46; F5: 43; F6: 46; Clava: 100. Forewing: 515:82; longest marginal seta: 182. Hind wing: 484:22; longest marginal seta: 145.
MALE. Length 430–515 µm. Similar to female but general body color darker (dorsum of mesosoma and metasoma brown to dark brown except anterior scutellum light brown and lobes of posterior scutellum yellow); appendages pale light brown except F2–F11 brown. Antenna as in Fig. 14 View FIGURES 13 – 15 . Forewing usually slightly wider than in female (6.2–6.5x as long as wide), with disc notably more setose and with a relatively smaller bare area, if any. Genitalia ( Fig. 15 View FIGURES 13 – 15 ) elongate, with hooked digiti.
Female diagnosis. This new species belongs to the incarnatus species group as defined by Chiappini et al. (1996). Females differ from A. daanei and A. tretiakovae by a relatively longer ovipositor, as indicated in the key, and from A. epos primarily by forewing proportions (6.3–6.7x as long as wide in A. vulneratus , 7.9– 8.6x as long as wide in A. epos ) and shorter ovipositor (2.5–2.7x as long as protibia in A. vulneratus , 2.8–3.1x as long as protibia in A. epos ), and also by F3 usually lacking a mps, although females of both species may occasionally have a mps on F3 of one antenna but lack one from the other. All these species are also genetically distinct (see below). Females of A. vulneratus differ from A. empoascae in usually lacking a mps on F3 and also in having a relatively longer ovipositor. Females of A. empoascae always bear a mps on F3 of both antennae, and the ovipositor is 2.2–2.4x as long as the protibia ( Triapitsyn 1997). Males of A. vulneratus are very similar to those of A. daanei and A. epos .
Etymology. The specific name is an adjective referring to the likely leafhopper host of this species.
Host. Erasmoneura vulnerata (Fitch) , most likely. Females of A. vulneratus [as Anagrus new species] from Grand Junction, Colorado, failed to parasitize fresh eggs of Homalodisca vitripennis (Germar) under quarantine laboratory conditions ( Morse & Stouthamer 2007).
Comments. Zimmerman et al. (1996) likely referred to this species as “ Anagrus epos Girault ”, but this is now impossible to verify because no voucher specimens of their study are available. Interestingly, and contrary to the earlier report by Zimmerman et al. (1996), we did not find any specimens of Erythroneura ziczac Walsh on grapevines in Grand Junction and Palisade, Mesa County, Colorado. The grapevines in the commercial and experimental vineyards there were predominantly infested by E. vulnerata , whereas the following deltocephaline leafhoppers were also present in much smaller numbers: Balclutha sp., Circulifer tenellus ( Baker) , and Colladonus sp.
Anagrus epos Girault : González et al. 1988: 23–25 (misidentification, in part [specimens from Sonora]). Anagrus (Anagrus) epos Girault : Triapitsyn 1998: 100–103 (misidentification, in part [specimens from Baja California and Sonora]).
Anagrus View in CoL sp.: Morse & Stouthamer 2007: 95.
Material examined. MEXICO, Sonora, W of Hermosillo (Carretera a Bahía Kino km. 12.6), Campo Experimental INIFAP-CECH "Costa de Hermosillo", 27.vii.1994, D. González, S.V. Triapitsyn, D. Powell, A. Fú-Castillo (from eggs of Erasmoneura variabilis (Beamer) on grape leaves, University of California, Riverside quarantine S&R # 94–33), 1 female, 1 male on slides and numerous females and males in ethanol [ UCRC]. Also 1 male from Baja California and numerous specimens of both sexes from Sonora listed by Triapitsyn (1998).
Host. Erasmoneura variabilis (Beamer) .
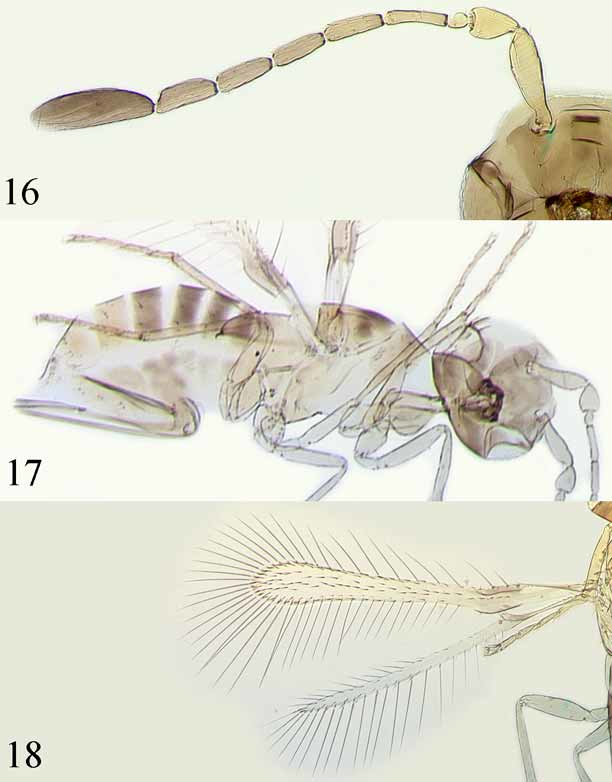
Comments. According to the molecular data presented below, specimens from Sonora from eggs of Erasmoneura variabilis on grape that were listed under A. epos by Triapitsyn (1998) likely belong to a separate species. Morphologically, females are quite similar to A. vulneratus from Colorado, including the antenna ( Fig. 16 View FIGURES 16 – 18 ), which often has a mps on F3, and the forewing ( Fig. 18 View FIGURES 16 – 18 ), which often lacks a bare area on the broadest part of the disc. The ovipositor ( Fig. 17 View FIGURES 16 – 18 ), however, is slightly shorter than in A. vulneratus from Colorado, and the external plate of the ovipositor has 3 distal setae. Since the Mexican specimens are genetically somewhat different from A. vulneratus from Colorado we prefer to call them A. sp. near A. vulneratus . As noted by Triapitsyn (1998), A. sp. near A. vulneratus (as A. epos from Sonora) is very similar morphologically to A. empoascae whose forewing, however, has a small, more or less defined bare area in the broadest part; also, the latter species parasitizes mainly eggs of Empoasca spp. on weeds and other low vegetation and crops ( Triapitsyn 1997, 2002). Their species status needs to be further investigated using molecular methods and cross-breeding experiments but that is beyond the scope of this study.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anagrus (Anagrus) epos Girault
| Triapitsyn, Serguei V., Rugman-Jones, Paul F., Jeong, Gilsang, Morse, Joseph G. & Stouthamer, Richard 2010 |
Anagrus
| Morse 2007: 95 |
Anagrus
| Morse 2007: 95 |
Anagrus epos
| Gonzalez 1988: 23 |