
Misgurnus amamianus, Nakajima & Hashiguchi, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5162.5.4 |
|
publication LSID |
lsid:zoobank.org:pub:02E91F18-BF2D-4A6C-859F-2D29D9F9DE94 |
|
DOI |
https://doi.org/10.5281/zenodo.6818072 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD8791-F018-897D-FF0E-FD81FE2BFF75 |
|
treatment provided by |
Plazi |
|
scientific name |
Misgurnus amamianus |
| status |
sp. nov. |
Misgurnus amamianus sp. nov.
( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 , 7a View FIGURE 7 )
Misgurnus anguillicaudatus (Cantor, 1842) View in CoL : Ogawa 1937: 18, Fig. 15 ( Japan: Tokuno-shima Is., Amami Isls., Kagoshima Pref.).
Misgurnus sp. IR (sensu Shmizu et al. 2011): Shimizu et al. 2011: 144, Fig. 1 View FIGURE 1 (OMNH-P20968, Japan: Iriomote-jima Is., Yaeyama Isls., Okinawa Pref.); Nakajima & Uchiyama 2017: 72, 74 ( Japan: Amami Isls., Kagoshima Pref.), 75 ( Japan: Iriomotejima Is., Yaeyama Isls., Okinawa Pref.); Nakajima 2018: 111 (no data); Hosoya 2019: 180 ( Japan: Kagoshima Pref.).
Holotype. KPM-NI 68404, male, 83.8 mm SL, Japan: Isen , Tokuno-shima Is., Amami Isls., Kagoshima Pref., 12. XII. 2015., T. Kitano leg.
Paratypes. 16 males and 10 females, 55.1–126.3 mm SL, all from the Ryukyu Islands , Japan. TKPM-P 26178, 26179, male & female, same data as the holotype; JNC353 , 354 , male & female, same locality as the holotype, 29. VII. 29. 2016., J. Nakajima leg.; KMNH VR 100463 , male, Noda, Isen, Tokuno-shima Is., Amami Isls. , Kagoshima Pref., 20. VIII. 2015., Y. Kano leg.; JNC554 , female, same locality, 10. XI. 2019., J. Nakajima leg.; NSMT-P 34733, male, Otsukan, China, Okinoerabu-jima Is., Amami Isls., Kagoshima Pref., 3. VIII. 1958., S. Ueno & Y. Morimoto leg.; OMNH-P 1145, 1146, male & female, Kamihirakawa, China, Okinoerabu-jima Is., Amami Isls. , Kagoshima Pref. , 29. VIII. 1964., Y. Shibata leg.; KPM-NI 68405, 68406, KMNH VR 100188, TKPM-P 26180, 26181, JNC140 , 141 , four males & three females, Kamihirakawa, China, Okinoerabu-jima Is., Amami Isls., Kagoshima Pref., 17. XI. 2013., J. Nakajima leg.; YCM-P 31002, male, Suitengu-no-ike, Kikai-jima Is., Amami Isls., Kagoshima Pref., data unknown (1930s)., H. Ikeda leg.; NSMT-P 14268, male & female, Amuro, Uken , Amami-oshima Is. , Amami Isls. , Kagoshima Pref. , 7. VIII. 1970., collector unknown; NSMT-P 28933 , two males & two females, Nishi-nakama, Amami-oshima Is., Amami Isls., Kagoshima Pref. , 4. IV. 1975., H. Sakai & M. Sato leg.; JNC128 , 1 male, Urauchi, Iriomote-jima Is., Yaeyama Isls., Okinawa Pref., 26. X. 2007., H. Mizutani leg.; OMNH-P 37946, male, same locality, 24. X. 2008., T. Suzuki, M. Hosokawa & M. Baba leg.; WMNH-PI 0311, male, Kumotta, Urauchi, Iriomote-jima Is., Yaeyama Isls., Okinawa Pref. , 8. IX. 2011., J. Nakajima leg.
Non-type specimens. OMNH-P 37941-37945, 37947, two males & four females, Japan: Urauchi , Iriomotejima Is., Yaeyama Isls., Okinawa Pref. 24. X. 2008., T. Suzuki, M. Hosokawa & M. Baba leg.
Diagnosis. The new species is distinguished from other species of Misgurnus by the combination of the following characteristics ( Table 3 View TABLE 3 ): lamina circularis at base of pectoral fin in adult male poleaxe-shape (vs. spatula-shape in M. buphoensis and M. nikolskyi ; elongated horizontally in M. dabryanus ; absent in M. fossilis ); neckline at base of lamina circularis shallow (vs. deep in M. anguillicaudatus ); dorsal fin ray iii+6 (vs. commonly iii+ 7–8 in M. anguillicaudatus ); last ray of dorsal fin commonly not branched (vs. commonly branched in M. anguillicaudatus , M. buohoensis , M. dabryanus , M. fossilis , M. mohoity and M. nikolskyi ); L/S less than 0.5 (vs. over 0.5 in M. anguillicaudatus , M. buphoensis , M. dabryanus , M. mohoity and M. nikolskyi ); LPC/DPA over 1.1 (vs. less than 1.0 in M. anguillicaudatus ( China clade), M. dabryanus and M. tonkinensis ); barbel length general (vs. long in M. dabryanus and M. tonkinensis ); caudal-peduncle depth shallow (vs. deep in M. dabryanus , M. multimaculatus and M. tonkinensis ); and total vertebrae number 44–48 (vs. over 49 in M. buphoensis , M. fossilis and M. nikolskyi ).
Description. The morphometric and meristic data for the type specimens (17 males and 10 females) are summarised in Table 2 View TABLE 2 . Data for the holotype are presented first, followed by those for the paratypes in parentheses, if different.
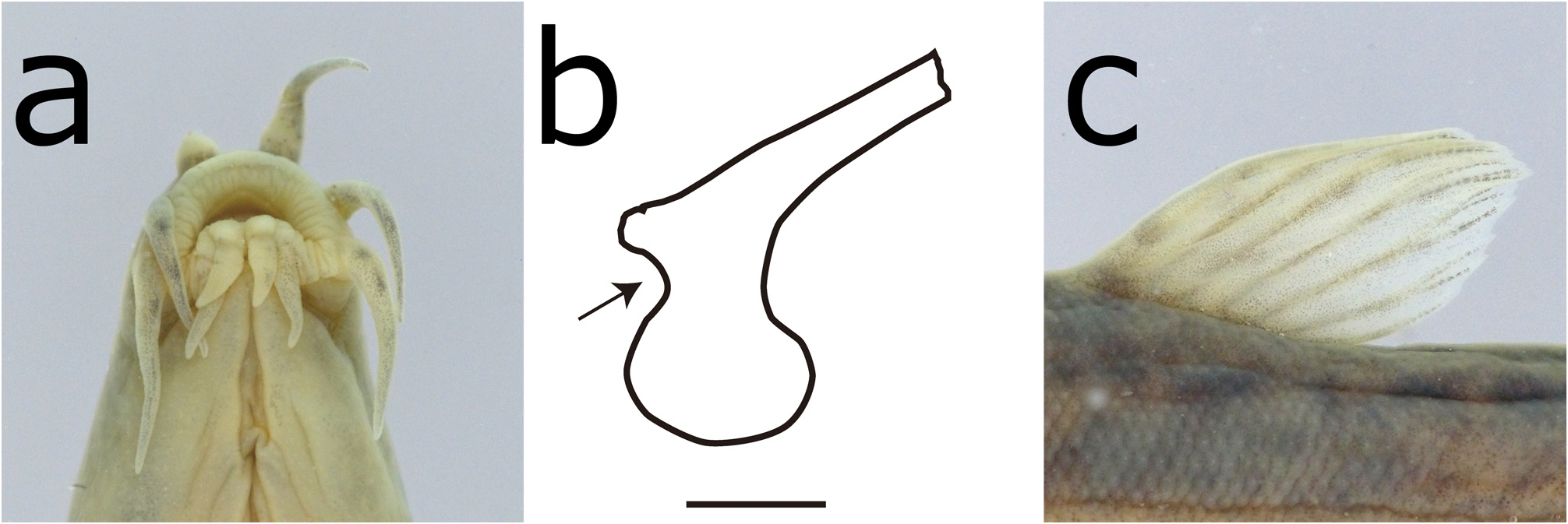
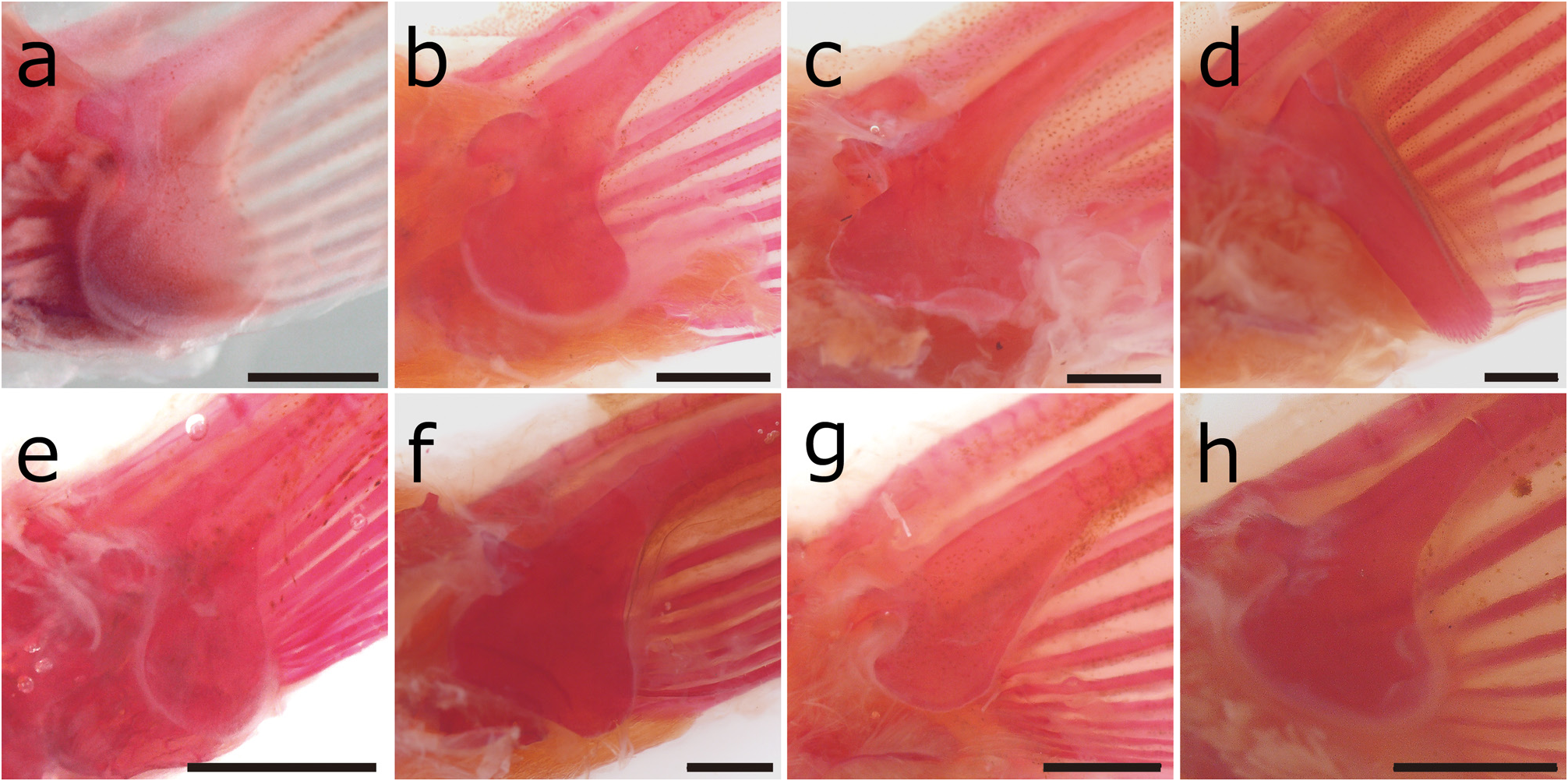
Body elongated, slightly compressed. Head elongated, snout rather short. Interorbital space convex. Eye moderately small, positioned upper and middle on head. Caudal peduncle compressed; ridges on the caudal peduncle at upper and lower edge weakly developed. Mouth small, inferior arched with fleshy lips; lower lip with 4 long, barbellike mental lobes; upper lip with transverse wrinkles on surface ( Fig. 4a View FIGURE 4 ). Barbels 3 pairs, first on rostrum, second on maxilla, and third on mandible; each barbels well developed, approximately 30% of head length. No lateral line. Ridges on caudal peduncle at upper and lower edge weakly developed. Small cycloid scales on trunk; head without scale. Lamina circularis at base of pectoral fin in adult males, poleaxe shape, backend round; neckline at base of lamina circularis shallow ( Figs. 4b View FIGURE 4 , 7a View FIGURE 7 ). First branched soft ray of pectoral fin longer than others, broad and slightly extended; pectoral fin in males longer than that in females. Dorsal-fin ray thin and weak; last ray unbranched, its length less than half of second branched soft ray ( Figs. 4c View FIGURE 4 , 5b View FIGURE 5 ). Margin of dorsal fin rounded, slightly pointed at center tip. Dorsal-fin base behind middle of body. Pelvic-fin origin below third or fourth branched dorsal-fin ray. Anal fin not reaching caudal-fin base. Margin of caudal fin rounded, slightly pointed at center tip. Abdominal vertebrae 30 (27–30); caudal vertebrae 18 (15–19); total vertebrae 48 (44–48) (n = 20) ( Fig 5a View FIGURE 5 ).
Colouration of fresh specimens. Head and back blackish brown with irregular and indistinct spots; slightly purplish on cheeks and body; abdomen yellowish brown ( Figs. 3 View FIGURE 3 , 5e View FIGURE 5 ). Unclear streak running from tip of snout to occiput, crossing to eye. Dorsal and caudal fins with irregular and indistinct spots. Upper spot at caudal-fin base ambiguous, size smaller than eye diameter. Ridges on caudal peduncle at upper and lower edge tinged with yellowish.
Sexual dimorphism. A lamina circularis of pectoral fin only in males; pectoral fin in males longer than that in females. Generally, body size of females larger than that of males.
Mitochondrial DNA sequence. Control region : LC672439 View Materials (Holotype); 12SrRNA: LC278291 View Materials .
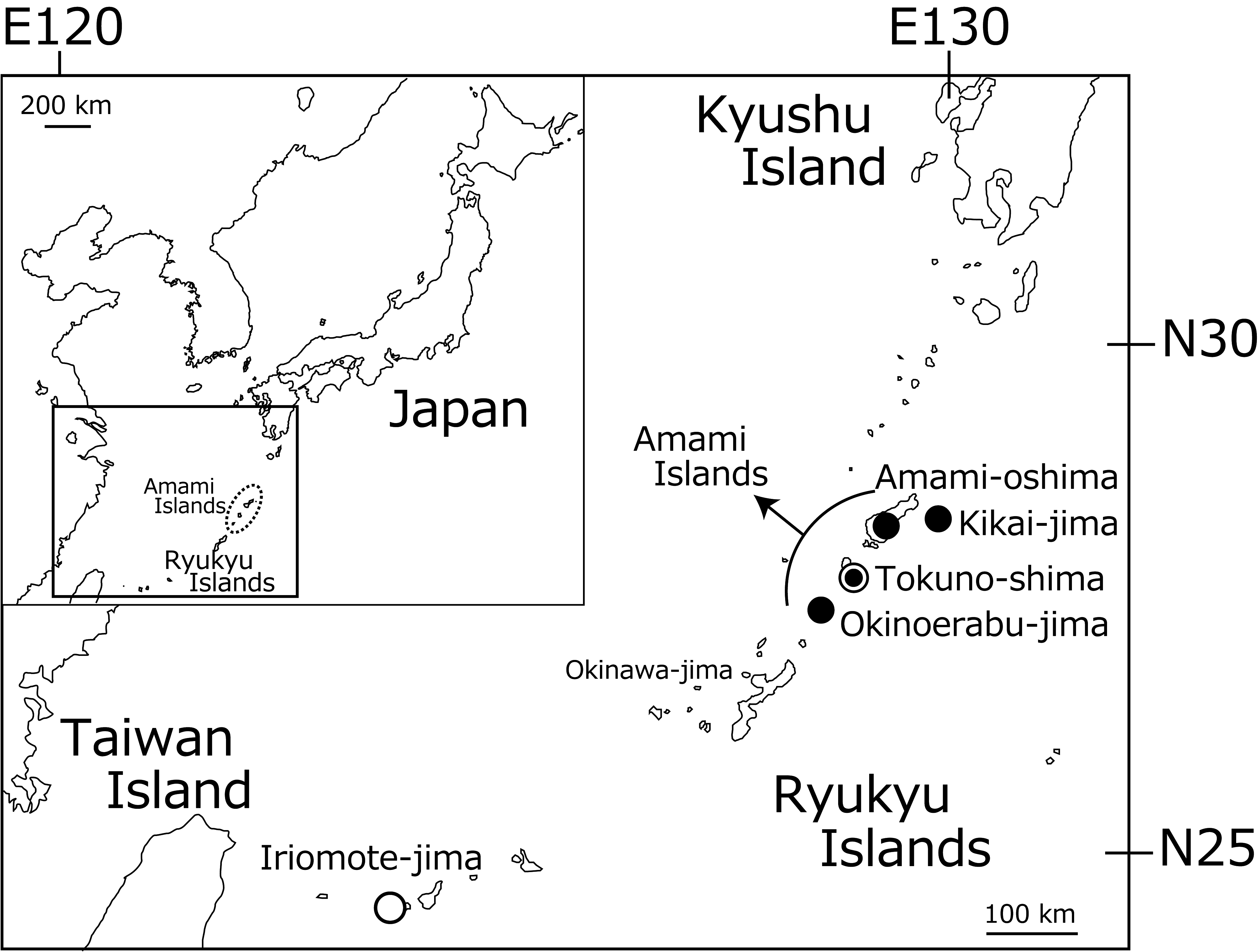
Distribution. Japan: Amami Islands. (Kikai-jima Island, Amami-oshima Isand, Tokuno-shima Island and Okinoerabu-jima Islamd), Kagoshima Prefecture, introduced to Iriomote-jima Island, Okinawa Prefecture, Japan ( Fig. 1 View FIGURE 1 ).
Habitat and biology. Misgurnus amamianus inhabits the muddy bottoms of small streams, paddy fields, and ponds with rich vegetation. The life cycle is unknown, but it breeds from May to August in captivity. The loach has a strong tendency to hide in vegetation.
Etymology. The specific name was derived from the Amami Islands, which is the primary distribution area of this species. Japanese name is suggested as Shinobi-dojyô.
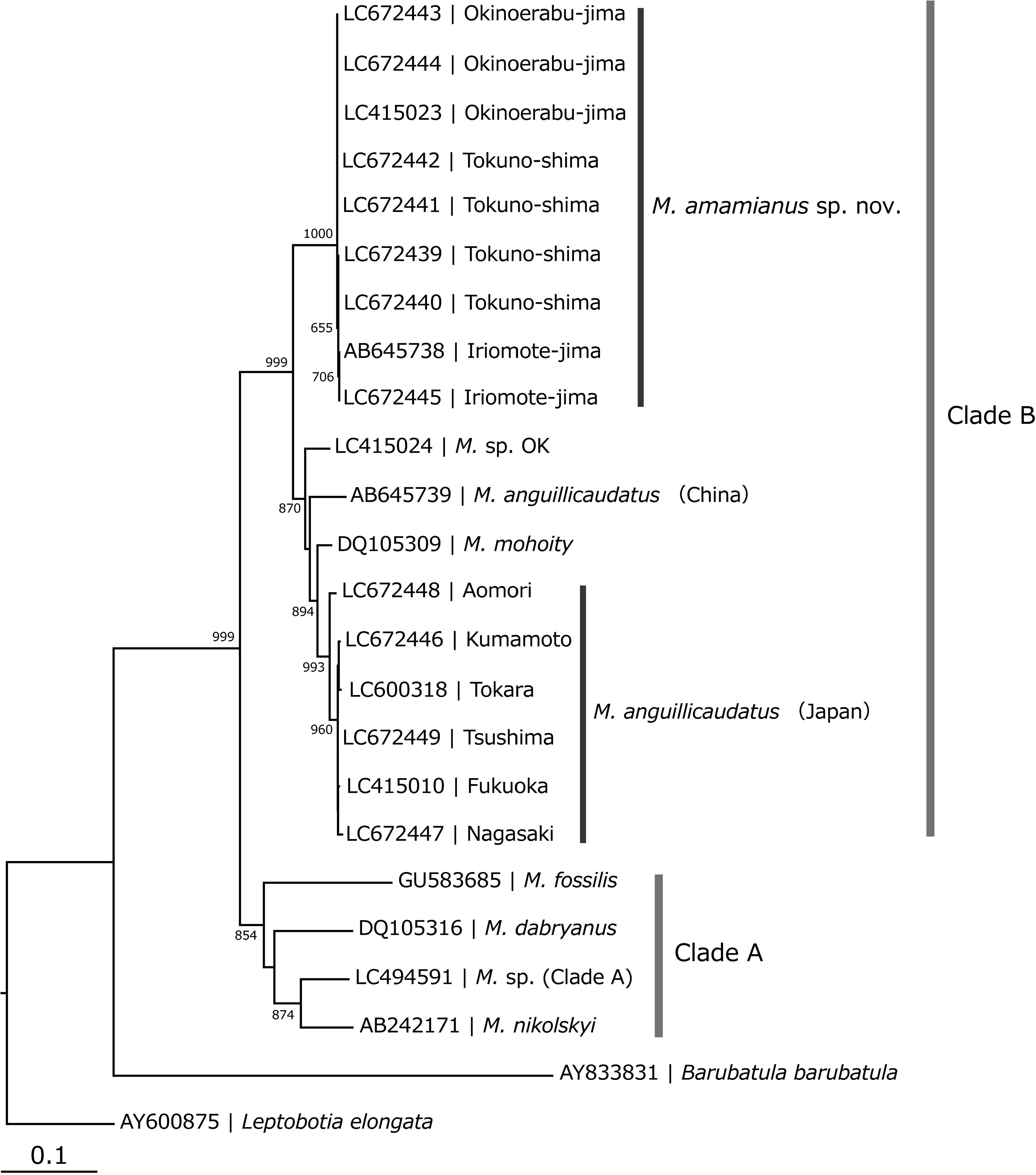
Remarks. There are many synonyms for Misgurnus anguillicaudatus , but no species or subspecies have been described from the Ryukyu Islands, including the Amami Islands ( Kottelat 2012). This species is morphologically and genetically distinguished from the known species and populations distributed in Northeast Asia ( Figs. 6 View FIGURE 6 , 7 View FIGURE 7 ; Table 3 View TABLE 3 ). In this study, we also compared the type specimens of Misgurnus anguillicaudatus as well as the five nominal species described from Japan and Taiwan Island. We confirmed that the combination of the ray number in the dorsal fin, the features of the last ray of the dorsal fin, the LPC/DPA ratio, the L/S ratio, and total vertebrae number morphologically can aid in the distinguishing of this species from M. amamianus ( Fig. 8 View FIGURE 8 ; Table 4 View TABLE 4 ).
There are two different mitochondrial DNA lineages in the genus Misgurnus : clades A and B (or Type I and Type II) ( Morishima et al. 2008; Koizumi et al. 2009; Kitagawa et al. 2011; Fujimoto et al. 2017; Okada et al. 2017). These two lineages were confirmed by phylogenetic analysis with high bootstrap values ( Fig. 6 View FIGURE 6 ). Phylogenetic analysis revealed that all M. amamianus individuals belonged to clade B and clearly diverged from other species within this clade ( Fig. 6 View FIGURE 6 ). The monophyly of M. amamianus is supported by a 100% bootstrap value ( Fig. 6 View FIGURE 6 ). Pairwise differences (i.e., p-distances) of the CR nucleotide sequences between M. amamianus and other Misgurnus species in clade B ranged from 6.3% to 7.8% (mean: 7.2%). These results support the validity of a distinct species, M. amamianus .
Many amphibian and reptile species are known to have evolved in this way in the central part of the Ryukyu Islands, including the Amami Islands (e.g., Lin et al. 2002; Matsui et al. 2005; Tominaga et al. 2013), and the same has been reported for freshwater fishes such as the Asian swamp eel, Monopterus ( Matsumoto et al. 2010) . Our phylogenetic analysis suggests that M. amamianus is also an endemic species that was isolated when the Amani Islands were separated from the Asian continent. Further detailed molecular phylogenetic analyses will help to clarify the evolutionary origins of this species.
Misgurnus amamianus is listed as Data Deficient (DD) in the Japanese Red List under the name Misgurnus sp. IR ( Ministry of the Environment 2020). Our survey showed that the last collection on Kikai-jima Island was in the 1930s, and the last collection on Amami-oshima Island was in the 1970s; the species has not been rediscovered since then. In addition, a pond that was the only habitat in Okinoerabu-jima Island (the collection site of the specimen used as the paratypes in this study) underwent renovation in the spring of 2021; the species may already be extinct. Therefore, there are only three definite habitats for this species: two ponds on Tokuno-shima Island and one paddy field on Iriomote-jima Island. In the Amami Islands, paddy fields which are the main habitat of this species, have been largely lost because of the rapid conversion from the once-common paddy rice cultivation to sugarcane cultivation ( Hagihara 1992). We believe that M. amamianus is on the verge of extinction, and urgent conservation measures are needed.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Misgurnus amamianus
| Nakajima, Jun & Hashiguchi, Yasuyuki 2022 |
Misgurnus anguillicaudatus (Cantor, 1842)
| Ogawa, K. 1937: 18 |