
Russula clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das, 2019
|
publication ID |
https://doi.org/ 10.5252/cryptogamie-mycologie2019v40a5 |
|
DOI |
https://doi.org/10.5281/zenodo.7825721 |
|
persistent identifier |
https://treatment.plazi.org/id/03DD3329-EF63-586F-FEE6-FF37C0DA59CB |
|
treatment provided by |
Felipe |
|
scientific name |
Russula clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das |
| status |
sp. nov. |
89. Russula clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das View in CoL View at ENA , sp. nov.
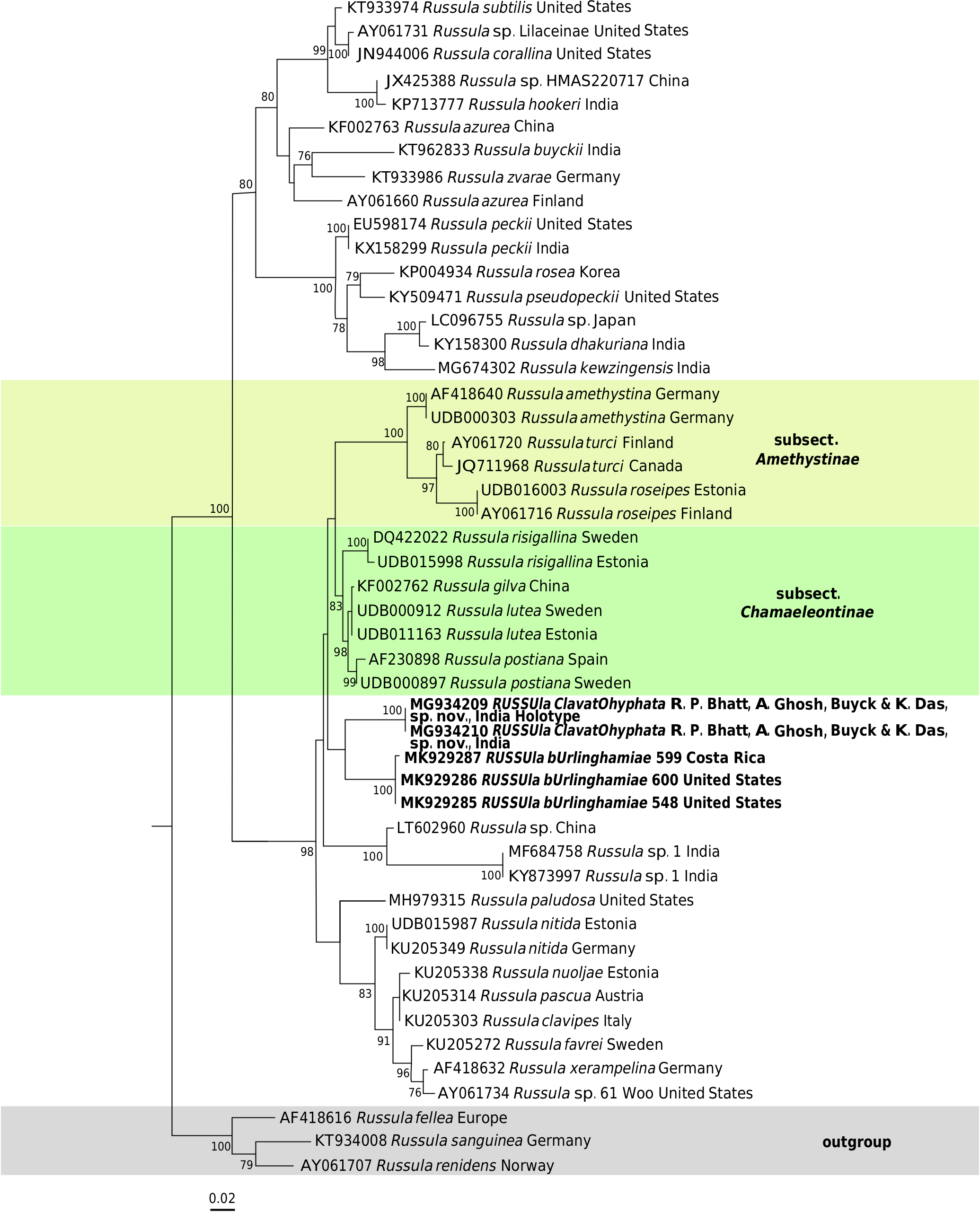
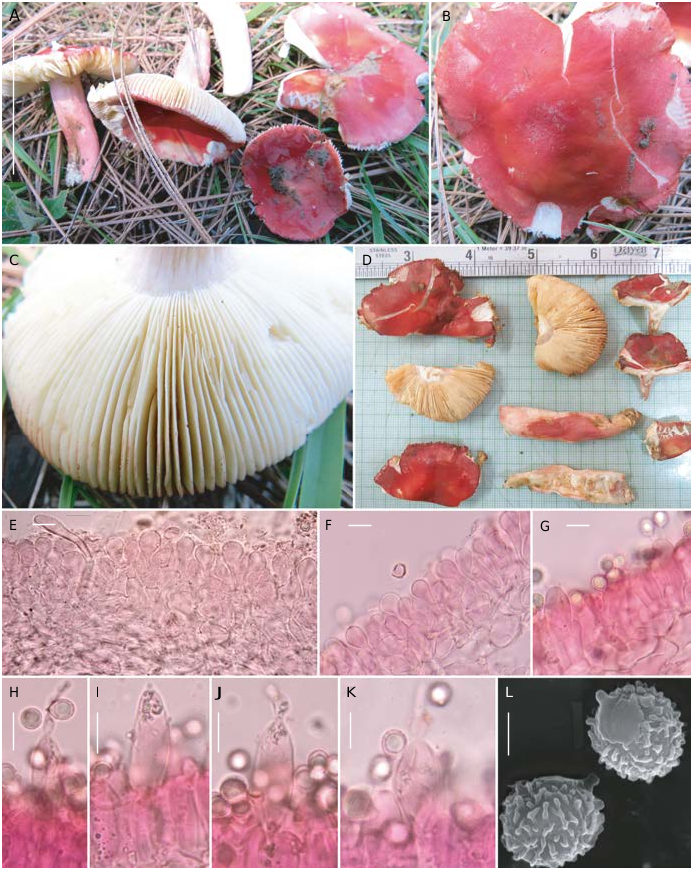
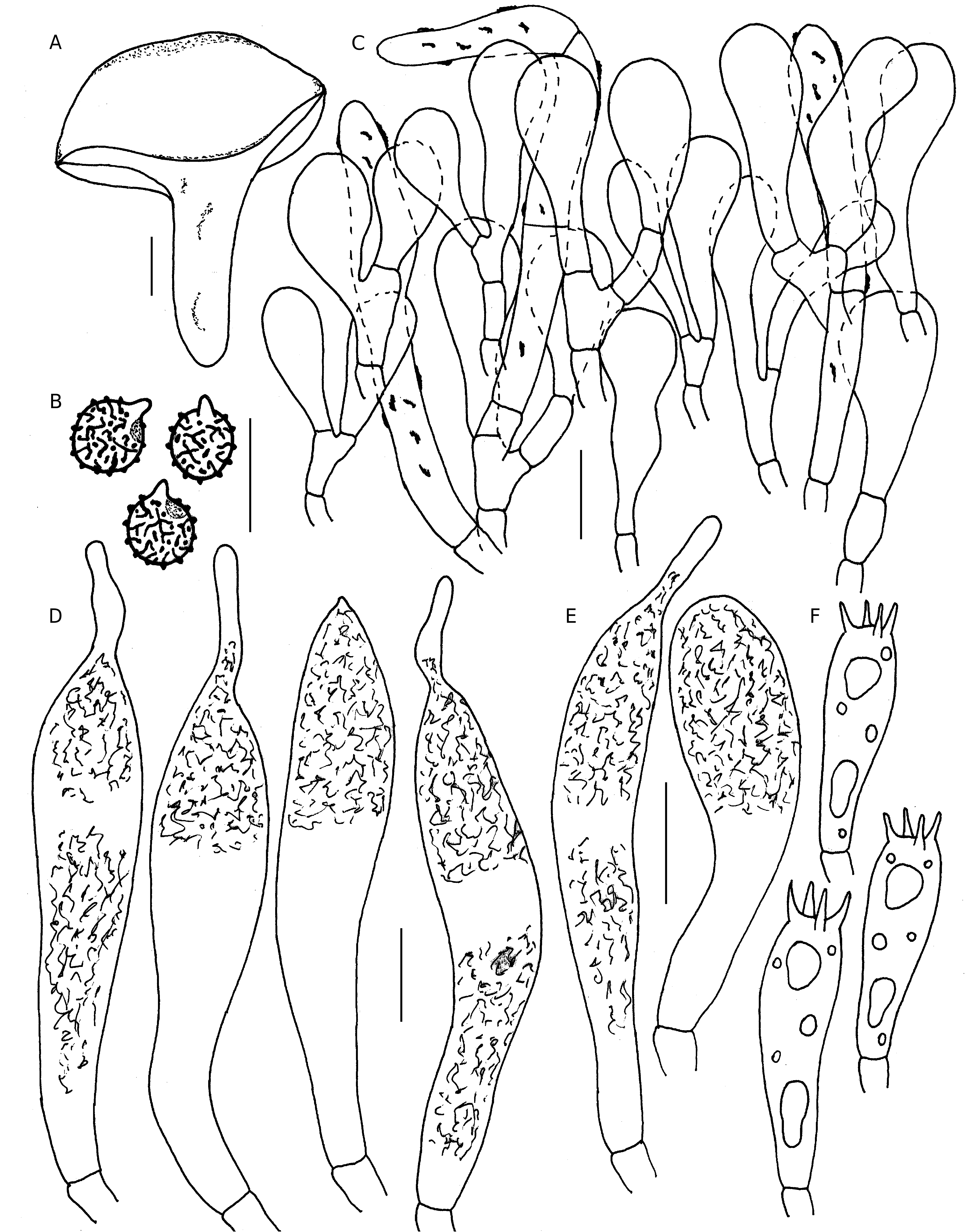
( Figs 19-21 View FIG View FIG View FIG )
Differs from R. luteobasis Peck essentially by the reddish pink coloration of the stipe and the slightly more ellipsoid spores, as well as because of its association with Pinus roxburghii Sarg.
TYPUS. — India. Utttarakhand, Rudraprayag district, Hariyali Devi forest, 30°15.955’N, 79°03.719’E, 1651 m asl., under Pinus roxburghii in temperate pure coniferous forest, 27.VII.2015, Aniket Ghosh, AG 15-756 (holo-, CAL [ CAL 1756 ). GoogleMaps
MYCOBANK. — 830765.
GENBANK. — MG934209 View Materials ( ITS holotype), MG934210 View Materials ( ITS paratype).
ETYMOLOGY. — referring to the clavate hyphal tips in pileipellis.
OTHER SPECIMENS EXAMINED. — Russula clavatohyphata . India. Rudraprayag district, Hariyali Devi forest, 30°15.952’N, 79°03.712’E, 1649 m asl., under Pinus roxburghii in temperate coniferous forest, 26.VII.2016, Aniket Ghosh, AG 16-1223 (para-, CAL[CAL 1757]). — Russula burlinghamiae . United States of America. Pennsylvania, Monte Alto, in mixed deciduous forest adjacent to University campus, 11.VIII.2005, 548/Buyck 05.108 (PC[PC0124708], GenBank MK929285 View Materials ); North Carolina, near Ashville, 17.VII.2004, 600/ Buyck 04.287 (PC[PC0124773], GenBank MK929286 View Materials ); Costa Rica. San José, Perez Zeledón, C.A.T.I.E. Experimental Forest of Villa Mills, 9.6989 -83.9419, 2850 m asl, in montane Quercus costaricensis forest, 12.VI.2001, leg. Buyck & Halling, 599/Buyck 01.237 (PC[PC0124772]f, GenBank MK929287 View Materials ).
DESCRIPTION
Basidiomata
30-62 mm in height.
Pileus
32-52 mm in diam., broadly convex when young, gradually planoconvex to applanate with depressed center, sometimes uplifted (towards margin) at maturity, sometimes broadly umbonate in the center; margin decurved to plane when mature, sometimes uplifted with maturity, entire, shortly tuberculate-striate; surface dry, viscid when moist, cracked to areolate with maturity, red to vivid red and paler towards the margin, peeling 1/2 of pileus radius, turning pale yellow to pastel yellow with 3% KOH.
Lamellae
Adnexed, subdistant (7-10 L/cm at pileus margin), yellowish white, sometimes forked near the stipe apex; edges marginate and red, at least near the pileus margin.
Lamellulae
Rare.
Stipe
53-58 × 6-14 mm, cylindric to subclavate, tapered at the base, central, dry, smooth, pale red or pastel pink to rose pink, tinged with ochraceous to ferruginous yellow near the base where the cuticle often cracks horizontally; stipe surface reacting straw yellow, wax yellow to mustard yellow and high red or vivid red with 3% KOH and guaiacol respectively.
Stipe context
Chalky white, stuffed to hollow with maturity; brownish orange to light brown after bruising or handling.
Taste
Mild.
Odour
Indistinctive.
Spore print
Pale yellowish white.
Basidiospores
(6.0-)6.34-6.98-7.63(-8.5) × (5.0-)5.53-6.11-6.7(-8.0) Μm, [n= 30, Q=(1.0-)1.05-1.15-1.24(-1.43)], subglobose to broadly ellipsoid, rarely ellipsoid; ornamentation amyloid (up to 0.5 Μm high), composed of conical to hemisphaerical warts mostly connected or aligned to give short ridges along with few isolated warts, either never forming incomplete reticulum or at most broken reticulum; suprahilar plage amyloid; apiculi up to 1.5 Μm high.
Basidia
22-54 × 9-13 Μm, 4-spored, subclavate to clavate; sterigmata up to 7 Μm long.
Pleurocystidia
47-73 × 9-11 Μm, subclavate to clavate, at apex appendiculate (appendages up to 30 Μm long), sub-moniliform, mucronate or rounded-obtuse, emergent up to 32 Μm beyond the basidiole tips, partly filled with heteromophous to granular or crystalline refringent content, SV-negative.
Lamella edges
Fertile with basidia and cystidia, and few cystidioid hyphal elements.
Cheilocystidia
35-60 × 7-10 Μm, subclavate to clavate with appendiculate, lageniform (appendages up to 11 Μm long) or rounded-obtuse apex with heteromophous and partly also filled with granular to crystalline refringent content, SV-negative.
Subhymenium layer
Up to 15 Μm thick, pseudoparenchymatous.
Hymenophoral trama
Mainly composed of large sphaerocytes and connecting hyphae.
Pileipellis
Up to 49 Μm thick, orthochromatic in cresyl blue, not sharply delimited from the underlying sphaerocytes of the context, trichoderm type, composed of erect to ascending, branched, septate, compact hyphal ends and with distinct primordial hyphae. Hyphal extremities near the cap margin composed of a single or a few cells only, terminal cells mostly bulbous, clavate, pyriform to subclavate with tapered base, measuring 9-34 × 5-11 Μm, with rounded or obtuse tips, subterminal cells cylindrical; in the cap center almost similar, mostly bulbous, clavate, pyriform to subclavate, measuring 10-37 × 6-12 Μm, with rounded or obtuse tips, subterminal cells cylindrical. Pileocystidia and cystidioid hyphae absent from all tissues. Primordial hyphae comparatively longer than the other extremities and often protruding or repent on the cap surface, occurring mostly singly, 4-7 Μm broad, thick-walled (up to 0.8 Μm), long, cylindrical, tapered at apex, septate (0-3 septa) with rounded, blunt or obtuse apex; incrustations present.
Clamp connections
Absent.
NOTES
In the field Russula clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das , sp. nov., is characterized by its red to vivid red coloured pileus and pale red or pastel pink to rose pink stipe. It may be confused with the unrelated R. sanguinea Fr. or R. rhodopus Zvára , two European species that are also ectomycorrhizal with coniferous trees, but both are moderately acrid or sometimes bitter, and differ further from the present taxon in their narrower, cylindrical pilear hyphae mixed with numerous pileocystidia with abundant contents ( Kränzlin 2005, Sarnari 2005, Knudsen et al. 2012).
The acystidiate pileipellis, amyloid suprahilar dot on the spores, mild taste, pale cream spore print and bulbous to clavate terminal cells mixed with dispersed, protruding primordial hyphae in pileipellis, undoubtedly place our species in the crown clade of Russula subg. Russula (sensu Buyck et al. 2018), near subsections Chamaeleontinae Singer and Amethystinae (Romagnesi) Bon. It differs from species in both latter subsections in the distinctly paler spore print, and from Chamaeleontinae additionally in its more robust and firm field habit.
Two species that are morphologically very similar to R. clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das , sp. nov., were described from America, but both are relatively unknown as they are apparently very rarely sporulating and have hardly been recollected. The first is R. burlinghamiae Singer , which principally differs from our species in the overall yellowish color and lack of any reddish tints. Since its original description, a modern, more detailed account of this rare North American species was published by Buyck et al. (2003) who also described a forma claricolor Buyck from the montane Quercus forests in Talamanca, Costa Rica. Russula burlinghamiae was placed with significant support in a clade comprised of Amethystinae and Chamaeleontinae in a recent multigene phylogeny ( Buyck et al. 2018). The second American species, R. luteobasis Peck , is even more similar to R. clavatohyphata, and even more rare than the previous one. It was in the past century only recollected in West Virginia (see mycoportal.org), but there are no reports or publications available for R. luteobasis since it was first described by Charles Peck (1904), except for the recently published type study ( Adamčík et al. 2018). Peck’s species differs from R. clavatohyphata R.P. Bhatt, A. Ghosh, Buyck & K. Das , sp. nov., essentially in the absence of a red color on the stipe surface and, most likely, also by its association with deciduous trees, but its major characters, such as spore ornamentation, composition and structure of pileipellis, spore print color, etc. are essentially the same.
Our ITS phylogeny retrieved no significant support to group R. burlinghamiae and R. clavatohyphata , nor did it obtain support to group either of these species with any other particular subsection (but neither did the multigene phylogeny by Buyck et al. 2018). Yet, it is almost certain that, once sequences will become available for R. luteobasis , the latter will turn out to be sister to the Indian species. The pale cream spore print of these American and Indian species adds support to their suggested basal position versus Amethystinae and Chamaeleontinae.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |