
Liotyphlops beui (Amaral, 1924)
|
publication ID |
https://doi.org/ 10.1016/j.jcz.2023.01.003 |
|
persistent identifier |
https://treatment.plazi.org/id/03DCB451-FF92-FFCF-B11E-FB96E3E5F89D |
|
treatment provided by |
Felipe |
|
scientific name |
Liotyphlops beui |
| status |
|
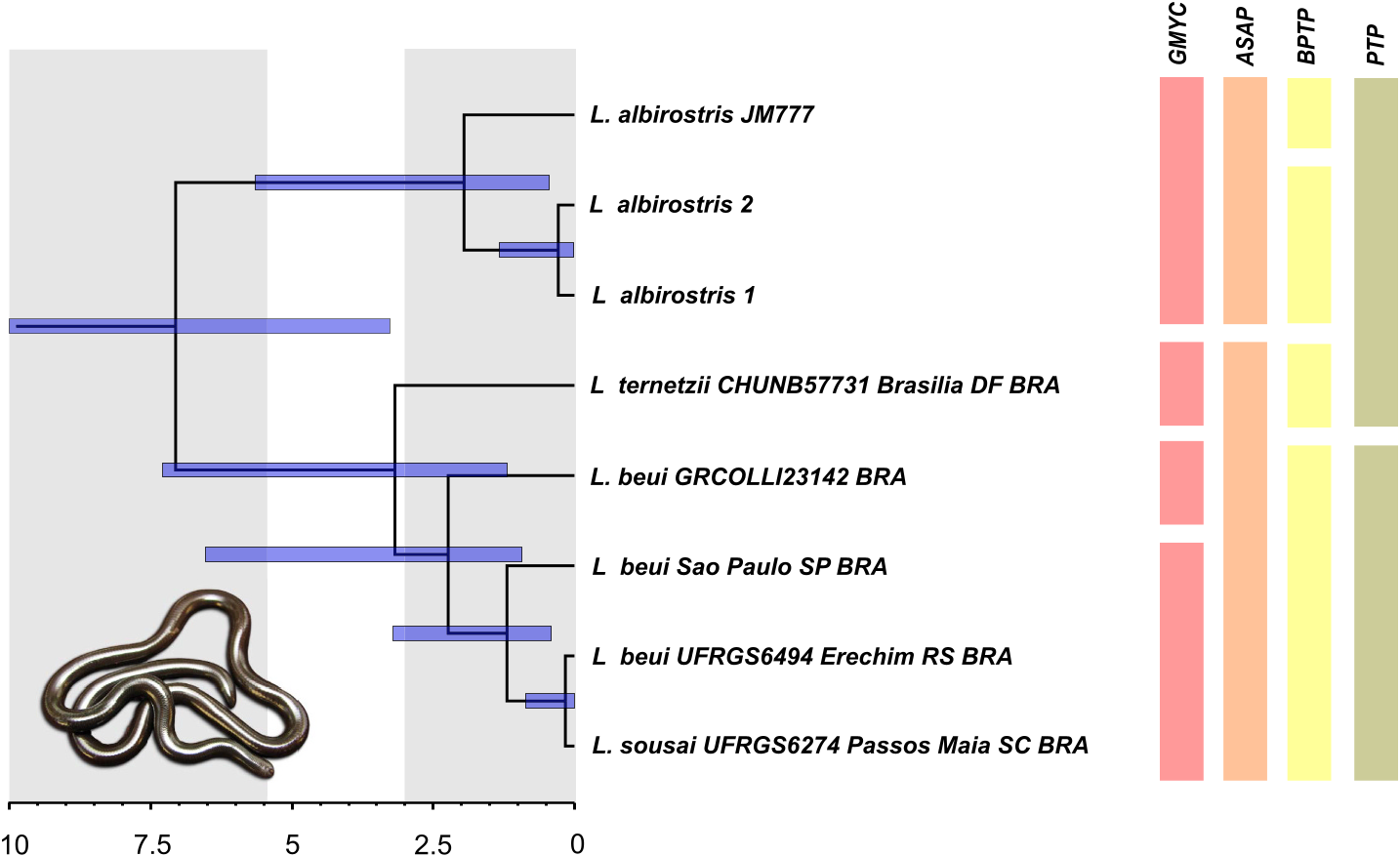
3.3. Redescription of L. beui View in CoL
L. beui ( Amaral, 1924) View in CoL
( Figs. 1 View Fig , 4–6 View Fig View Fig View Fig , 9–12 View Fig View Fig View Fig View Fig )
3.3.1. Synonymy
Helmintophis beui Amaral, 1924:25 . Holotype: IBSP 1806 View Materials (lost), from Instituto Butantan (761 m above sea level), S˜ao Paulo municipality, S˜ao Paulo state, Brazil. Paratypes: IBSP 281 View Materials (lost), 282 (Lost), 652 (lost), 1041 (lost), MCZ 16702 About MCZ , 17842 About MCZ , BMNH 1946.1 .11.12, also from Instituto Butantan , S˜ao Paulo municipality, S˜ao Paulo state, Brazil .
L. sousai Santos & Reis, 2018:507 View in CoL . Holotype: UFRGS 6274 View Materials (juvenile female), from Usina Hidrel´etrica Passos Maia (800 m above sea level), Passos Maia municipality, Santa Catarina state, Brazil. [New synonymy].
3.3.2. Heterochresonymy L. ternetzii View in CoL [non Helmintophis ternetzii Boulenger, 1896:584 ]: Amaral,
1924; Santos & Reis, 2018 (In error).
3.3.3. Diagnosis
This species can be diagnosed based on the following combination of characters: (1) anterior dorsal scale rows 22–24, usually 22; (2) midbody dorsal scale rows 20–22, usually 20; (3) posterior dorsal scale rows 20; (4) dorsal scales in vertebral row 395–446 (in males 395–420, in females 410–446); (5) mid-ventral scales 375–434 (in males 375–402, in females 395–434); (6) subcaudal scale rows 10–20 (in males 16–20, in females 10–15); (7) supralabials four; (8) infralabials three; (9) postfrontal scales contacting prefrontal three; (10) two scales in contact with posterior edge of nasal scale; (11) dorsal coloration uniformly dark brown or black, head and up to three first dorsal scale rows pinkish; (12) ventral coloration uniformly dark brown or black, head, gular region, and infralabials white or pink, cloacal region and ventral surface of tail white; (13) SVL 173–294 mm (in males 173–275 mm, in females 191–294 mm); (14) Tail length 4.2–12.5 mm (in males 6.7–12.5 mm, in females 4.2–6.6 mm).
3.3.4. Comparisons
Characters for other species are contained within parentheses, and our data has been supplemented with those provided by Dixon and Kofron (1983). L. beui is likely to be confused with another similar congener, L. ternetzii , that also has a black dorsal coloration and is widely distributed from Uruguay and Argentina to northern Brazil. There appears to be no difference among these species in skull morphology (O.M. Entiauspe-Neto, pers. obs.). While it has been suggested that both species could be distinguished based on head coloration, which should be pink or white in L. beui and black in L. ternetzii , an analysis of both types (BMNH 1946.1.11.77 - holotype of Helminthophis ternetzii ; BMNH 1946.1.11.12 - paratype of H. beui ) reveals that both specimens present a similar, white head coloration in preservative, with a distinctly colored light brown body. Until an integrative revision incorporating molecular data is made, the taxonomic status of these two species is only weakly supported through external morphology. In light of this, L. beui can be distinguished from L. ternetzii by having posterior dorsal rows 20 (22 in L. ternetzii ) and dorsal scales in vertebral row 395–446 (463–510 in L. ternetzii ) ( Dixon and Kofron, 1983; Centeno et al., 2010). However, it should also be noted that different values for these counts in literature can be attributed to possible misidentifications (e.g. reports of posterior dorsal rows 22 in L. beui by Santos & Reis, 2018).
Furthermore, L. beui can be distinguished from L. schubarti Vanzolini, 1948 by having the nasal scale in contact with the second supralabial (nasal separated from second supralabial by an accessory scale in L. schubarti ). It also differs from L. albirostris ( Peters, 1858) and L. wilderi ( Garman, 1883) by having a single scale contacting the posterior edge of nasal, between second supralabial and prefrontal (two scales contacting posterior edge of nasal, between second supralabial and prefrontal in L. albirostris and L. wilderi ). It furthermore differs from L. wilderi by having dorsal scales in vertebral row 395–446 (dorsal scales in vertebral row 304–358 in L. wilderi ). From other two species, L. anops ( Cope, 1864) and L. argaleus Dixon and Kofron, 1983 , L. beui can be distinguished by having dorsal scales in vertebral row 395–446 and usually three postfrontal scales contacting prefrontal (four postfrontal scales contacting prefrontal, posterior dorsal scale rows 22–24 in L. anops and L. argaleus ). Other congener which bears great resemblance to L. beui is L. caissara Centeno, Sawaya, & Germano, 2010 , from which it can be separated by having a black venter and two scales in contact with the posterior edge of nasal between second supralabial and prefrontal (white venter, one scale in contact with the posterior edge of nasal between second supralabial and prefrontal in L. caissara ). It can be distinguished from another congener which occurs in the northernmost range of the Atlantic Forest, L. trefauti Freire, Caramaschi, & Argolo ˆ, 2007, by having dorsal scales in vertebral row 395–446, a uniformly black dorsal and ventral coloration, and usually three postfrontal scales contacting prefrontal (four postfrontal scales contacting prefrontal, dorsal scales in vertebral row 520–543, uniformly brown dorsal and ventral coloration, four postfrontal scales contacting prefrontal in L. trefauti ). Lastly, it differs from L. taylori Santos & Reis, 2018 , by having three infralabials (two infralabials in L. taylori ).
3.3.5. Redescription
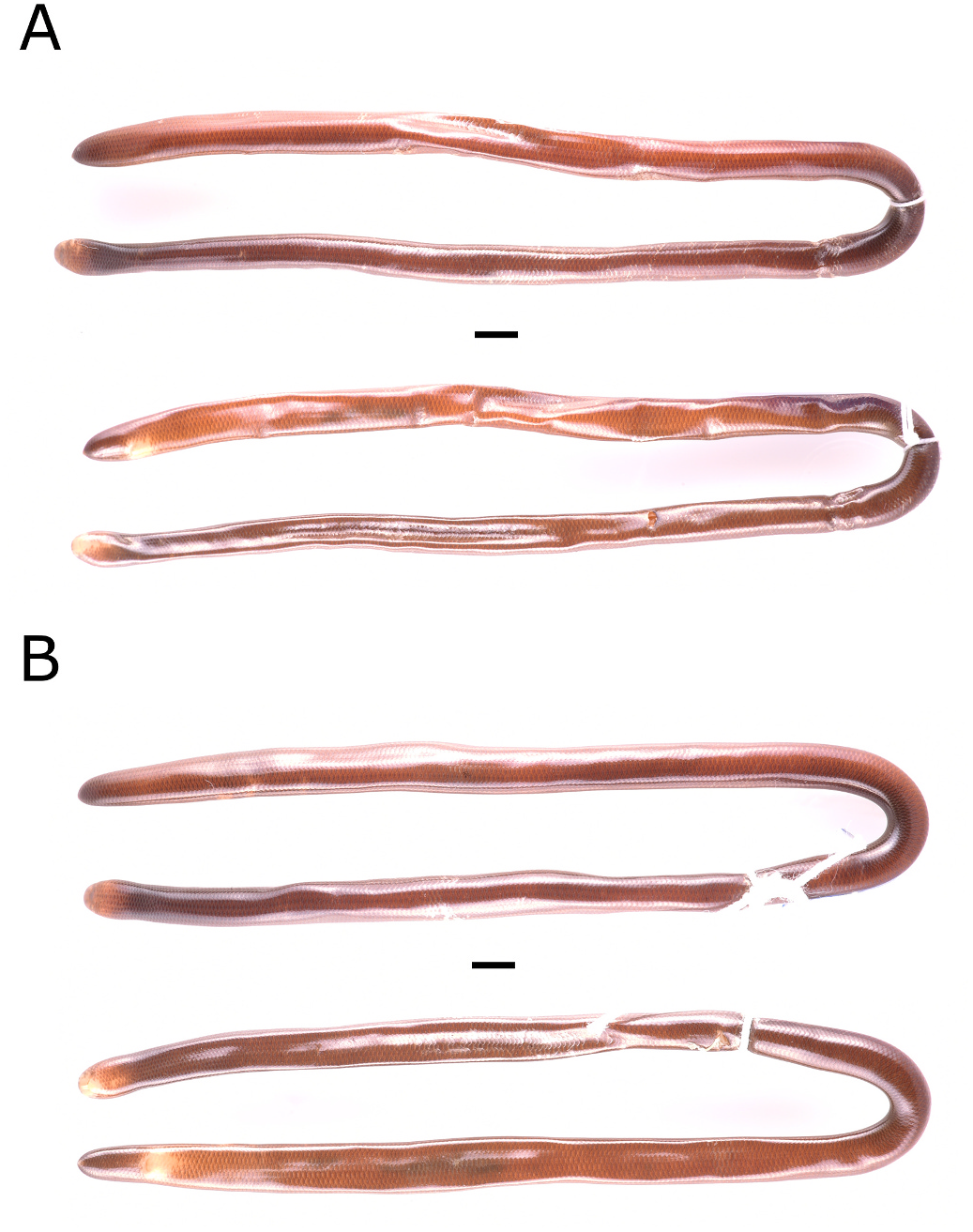
Redescription is based on a topotypical specimen (IBSP 90395,
Fig. 7A–B View Fig ), an adult female, from Instituto Butantan, S˜ao Paulo municipality, S˜ao Paulo state, Brazil. Snout-vent length 258 mm, head length 5.5 mm (2.13% of SVL), tail length 8 mm (3.10% of SVL). Rostral large, longer than wider, contacting nasals anterolaterally, prefrontals laterally, and frontal posteriorly; prefrontals paired, triangular, contacting rostral anterolaterally, nasal ventrally, and frontal dimorphic characters were found to be correlated ( Fig. 8 View Fig ). Total length ranges from 167 to 300 mm (245 ± 35, n = 28), in males ranges from 167 to 272 mm (232 ±30, n = 17), in females ranges from 186 to 300 mm (265 ±33, n =11). Snout-vent length ranges from 173 to 294 mm (249 ±32, n =28), in males ranges from 173 to 275 mm (241 ±31, n = 17), in females ranges from 191 to 294 mm (260 ± 30, n = 11). Tail length ranges from 4.2 to 12.5 mm (7.5 ±2.2, n =28), in males ranges from 6.7 to 12.5 mm (8.9 ±1.7, n = 17), in females ranges from 4.2 to 6.6 mm (5.3 ±0.7, n =11). Head length ranges from 3.4 to 5.6 mm (4.7 ±0.6, n =28), in males ranges from 3.5 to 5.6 mm (4.8 ±0.6, n =17), in females ranges from 3.4 to 5.5 mm (4.5 ± 0.6, n = 11). First row of dorsal scales range from 22 to 24 (22 ±0.8, n =28), and the last row of dorsal scales are 20 (n = 28). Postfrontal scales in contact with the frontal scale are three (n =28). Scales in contact with the nasal are two (n =28). Supralabial scales are four (n =28), infralabial scales are three (n = 28). Dorsal scales in vertebral row range from 395 to 446 (415 ± 16, n = 28), in males range from 395 to 420 (404 ± 6.2, n = 17), in females range from 410 to 446 (433 ± 10, n = 11). Mid-ventral scales range from 375 to 434 (399 ±18, n = 28), in males range from 375 to 402 (386 ±6, n =17), in females range from 395 to 434 (420 ±11, n = 11). Subcaudal scales range from 10 to 20 (16 ± 3, n = 28), in males range from 16 to 20 (18 ±1, n =17), in females range from 10 to 15 (12 ±1, n = 11). Head coloration in all specimens has white pigmentation (n = 28), ranging from fully (up to three dorsals) to mostly (up to one dorsal, or not reaching dorsals) white.
dorsoposteriorly; posterior edge of both prefrontals not contacting posterior edge of rostral; nasal plate divided, contacting rostral anteriorly, prefrontal dorsally, and supralabials 1–2 ventrally; eyespot inconspicuous, not clearly visible; supralabials 3/3; postfrontals five, three contacting prefrontal; infralabials 4/4; dorsal scale rows 24/20/ 20, smooth, cycloid; vertebral scales 394; midventral scales 353; subcaudal scales 14.
3.3.6. Coloration
In life, specimens have a uniformly dark brown or black dorsal coloration, with a pinkish or beige head coloration, that extends up to the first five dorsal scale rows. Ventral coloration uniformly dark brown, except for cloacal region, subcaudals, gular region, and infralabials that are white or pink. Body scales have a light brown outline on bottom and dark brown or black coloration on apex. In preservative, pink coloration becomes white, and black becomes light brown.
3.3.7. External morphology variation
We detected sexual dimorphism, in which females of L. beui attain higher total length (t 17,11 =2.7122, P <0.05, n =28), smaller tail length (t 17,11 = 6.2525, P <0.0001, n = 28), lower dorsal scales in vertebral row (U 17,11 = 3, Z = 4.2, P <0.00001, n = 28), lower mid-ventral scales (U 17,11 = 1, Z = 4.3, P <0.00001, n = 28), and lower subcaudal scales (U 17,11 = 0, Z = 4.3, P <0.00001, n = 28). All sexually 3.3.8. Geographic distribution
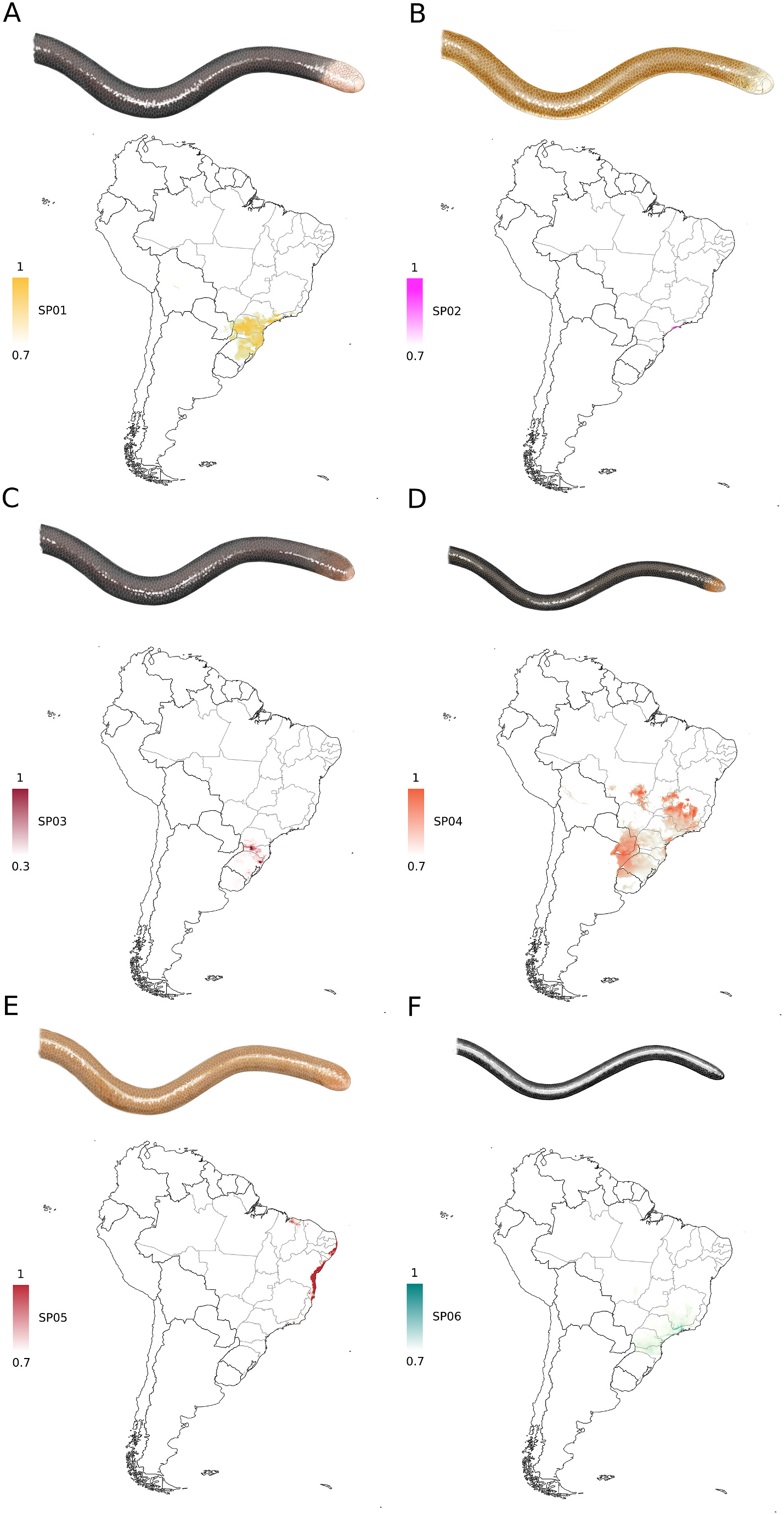
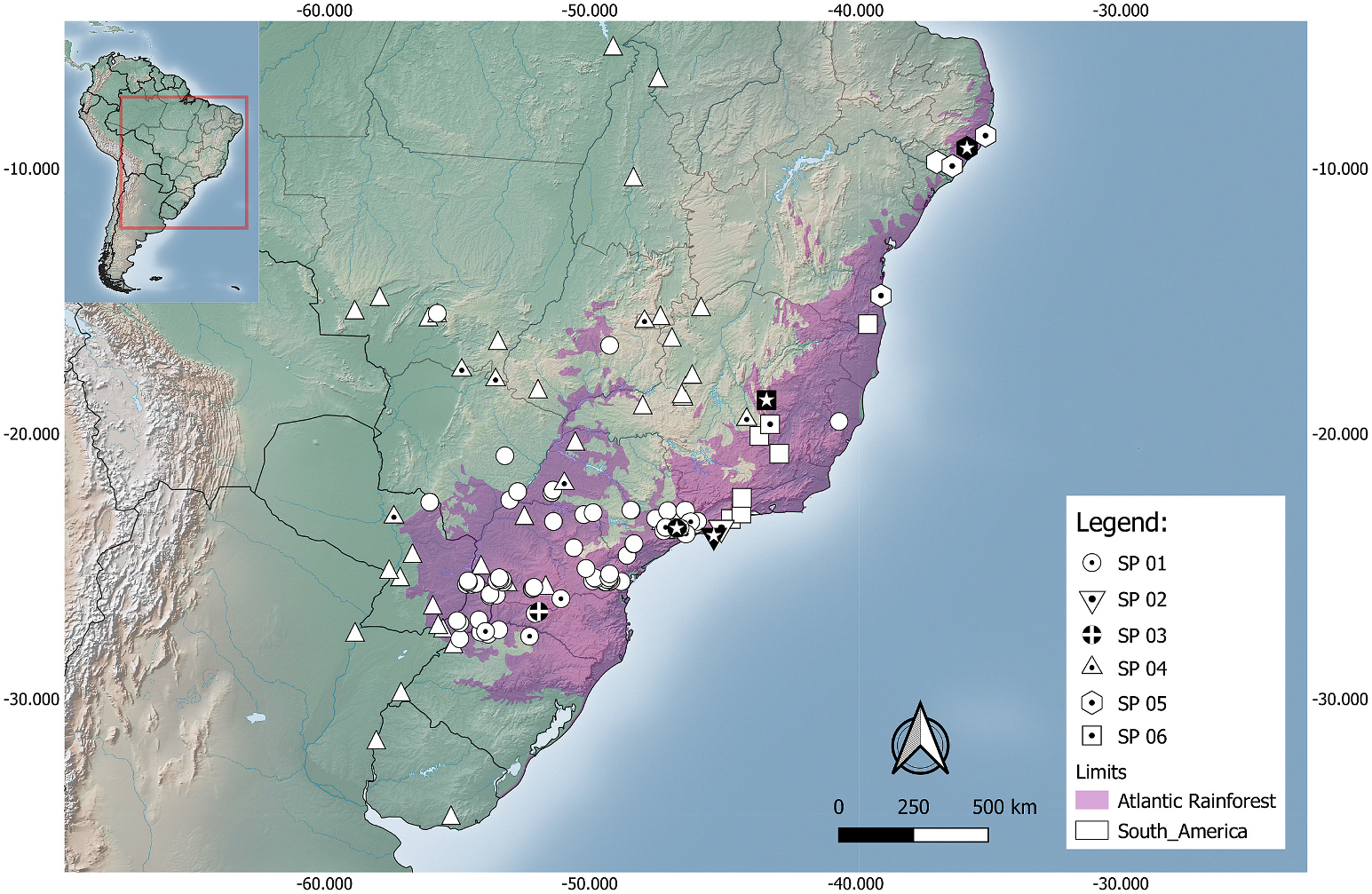
L. beui View in CoL has been recorded from Argentina, Brazil, and Paraguay ( Fig. 9 View Fig ), mostly from Atlantic Rainforest, although there have been extralimital records from the Cerrado, Pantanal, and Chiquitano Dry Forest, at low to moderate altitudes (up to 1000 m above sea level) ( Dixon and Kofron, 1983; Nogueira et al., 2019). Records of L. beui View in CoL are largely concentrated in southeastern and southern Brazil, in the states of S˜ao Paulo, Paran´a, and Rio Grande do Sul. Isolated records from its northernmost range, in the Cerrado of Mato Grosso and Goi´as states in Brazil, should be carefully reviewed regarding their identity and provenance, as these are separated from its core distribution in southeastern Brazil by at least 500 km. This species has an estimated Extent of Occurrence (EOO) of 1,295,329,935 km 2.
In its core geographic distribution, L. beui View in CoL is sympatric with L. ternetzii View in CoL in northwestern Rio Grande do Sul, western Parana ´, and western S˜ao Paulo. The latter species appears to be largely associated with the Cerrado and Chaco Dry Diagonal open areas, widely distributed from Para´, in northern Brazil, southwards into Uruguay. Other two congeners which L. beui View in CoL might be sympatric with in the Atlantic Rainforest are L. caissara View in CoL , which occurs in the coastal lowlands and marine islands of S˜ao Paulo, and L. wilderi View in CoL , which occurs in coastal lowlands and montane forests of Rio de Janeiro and Minas Gerais. Another congener, L. trefauti View in CoL , is recorded from the northernmost portion of the Atlantic Rainforest, in northeastern Brazil, where L. beui View in CoL has not been recorded.
3.3.9. Osteological comparisons
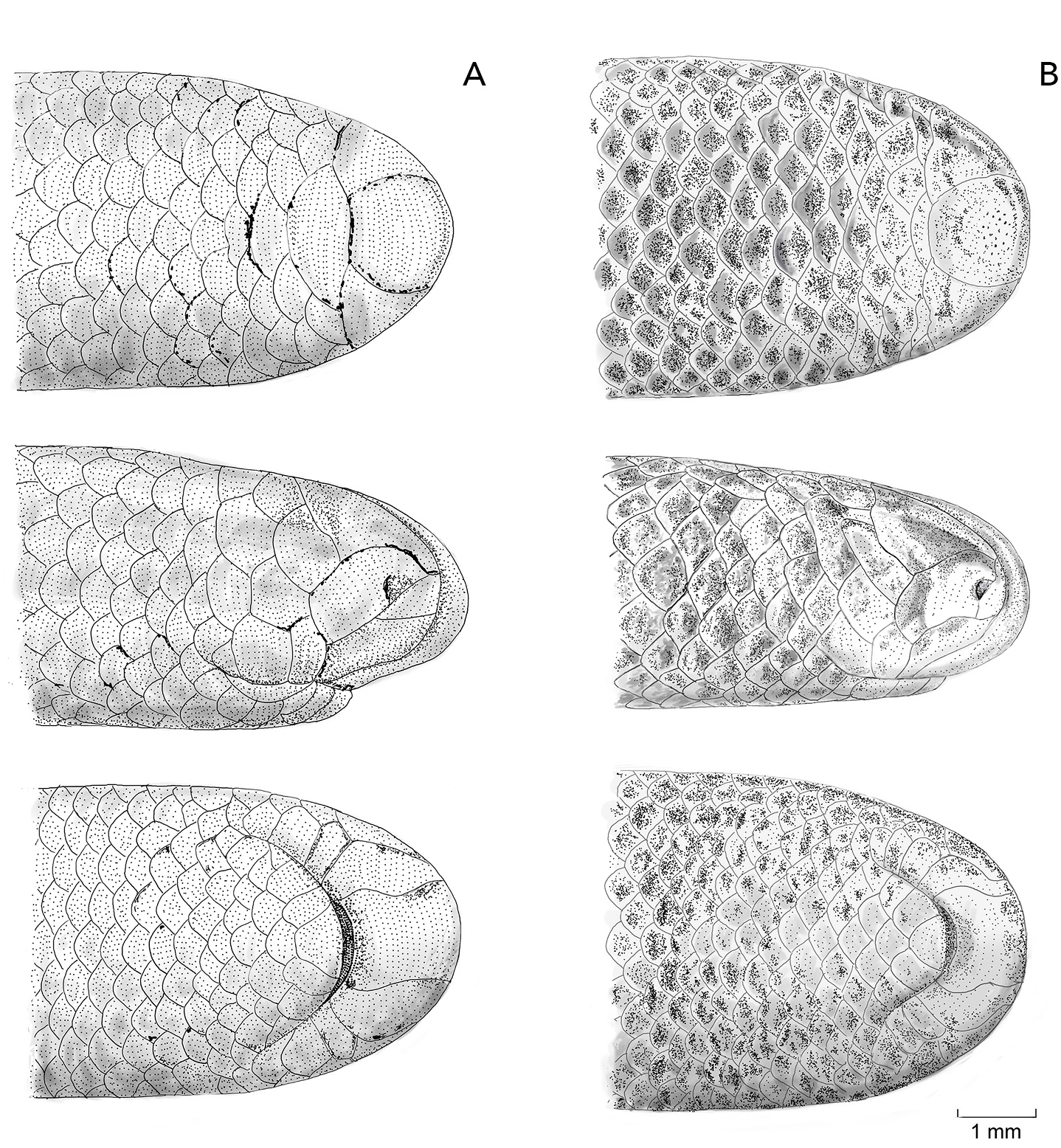
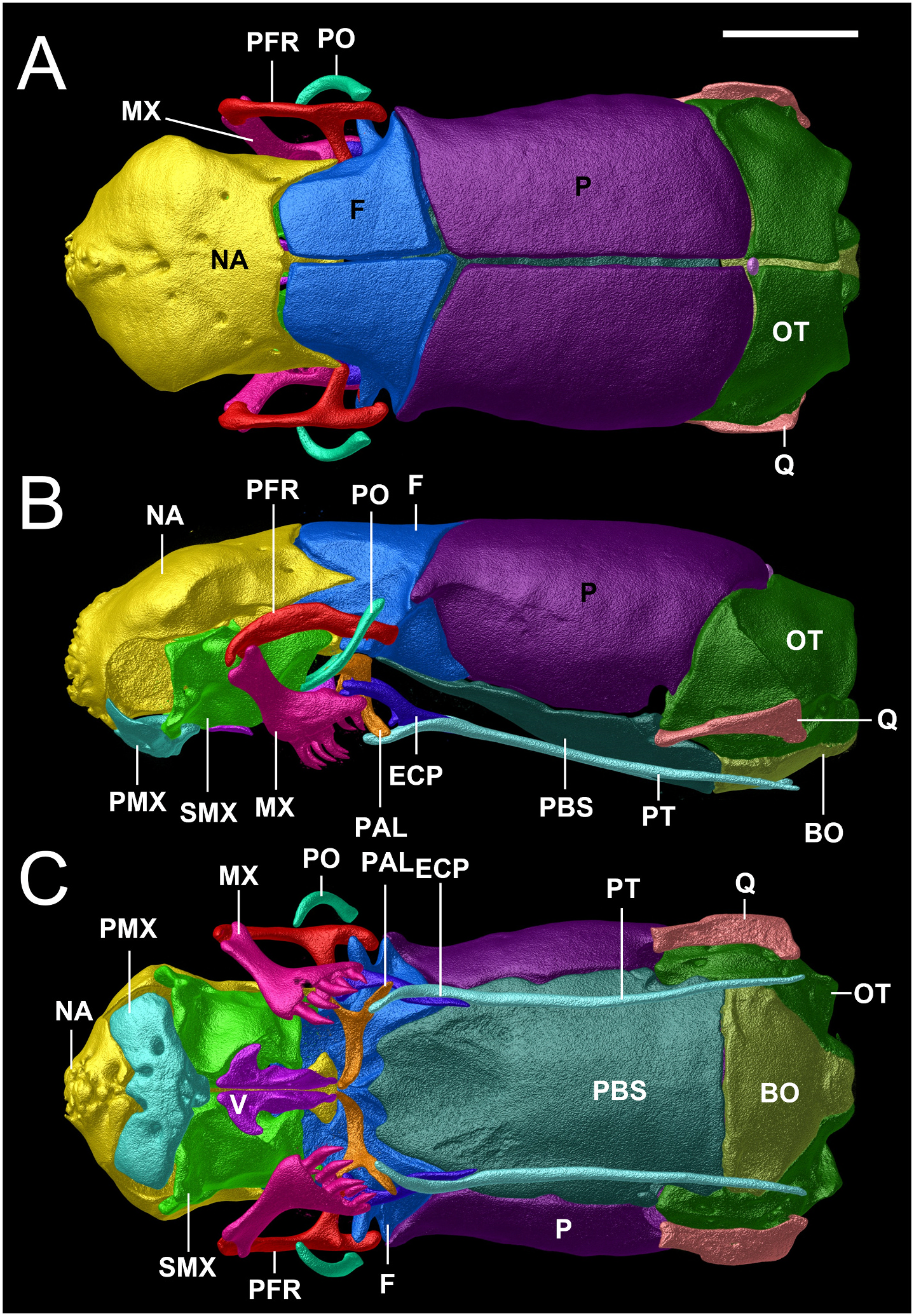
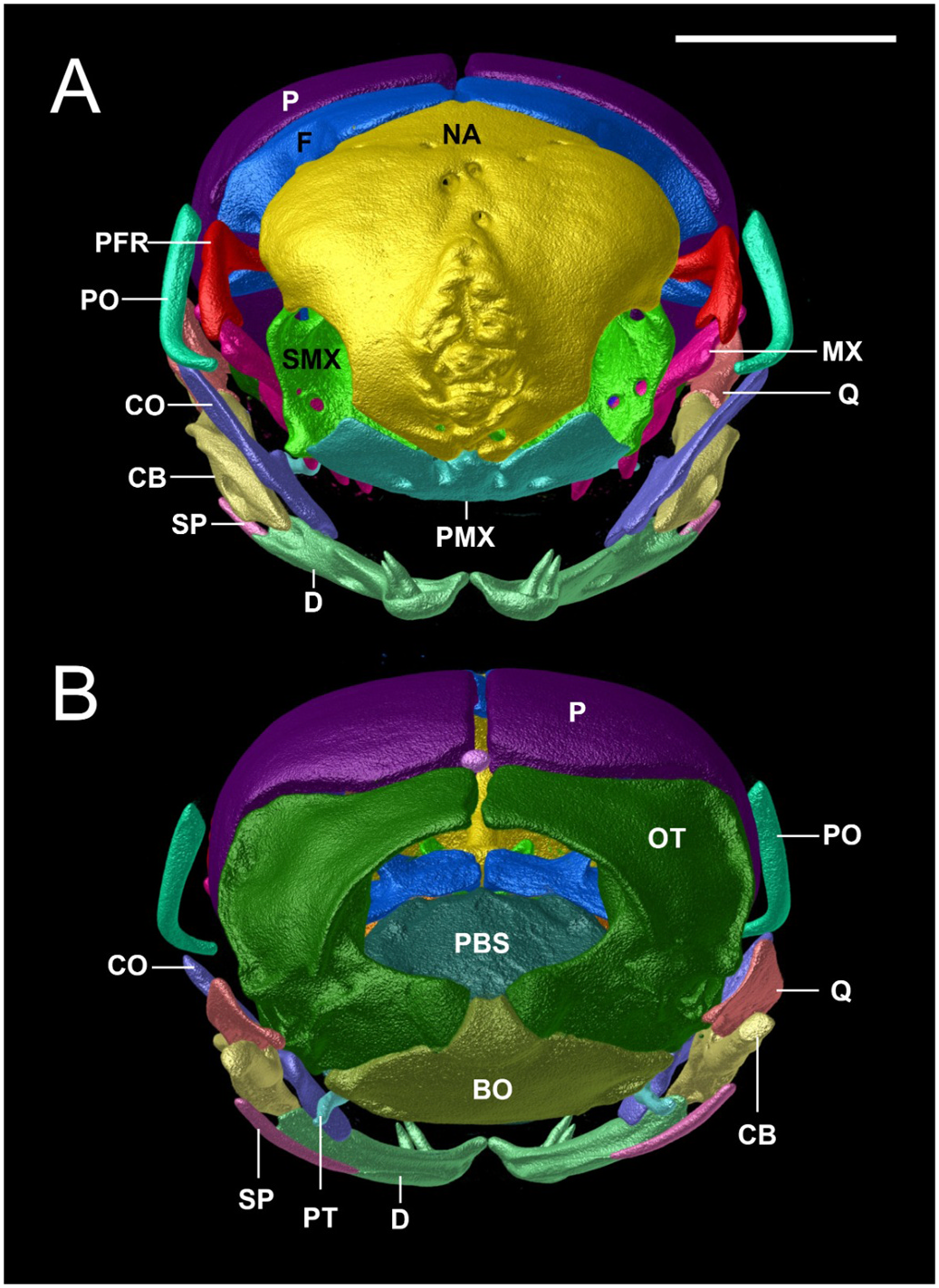
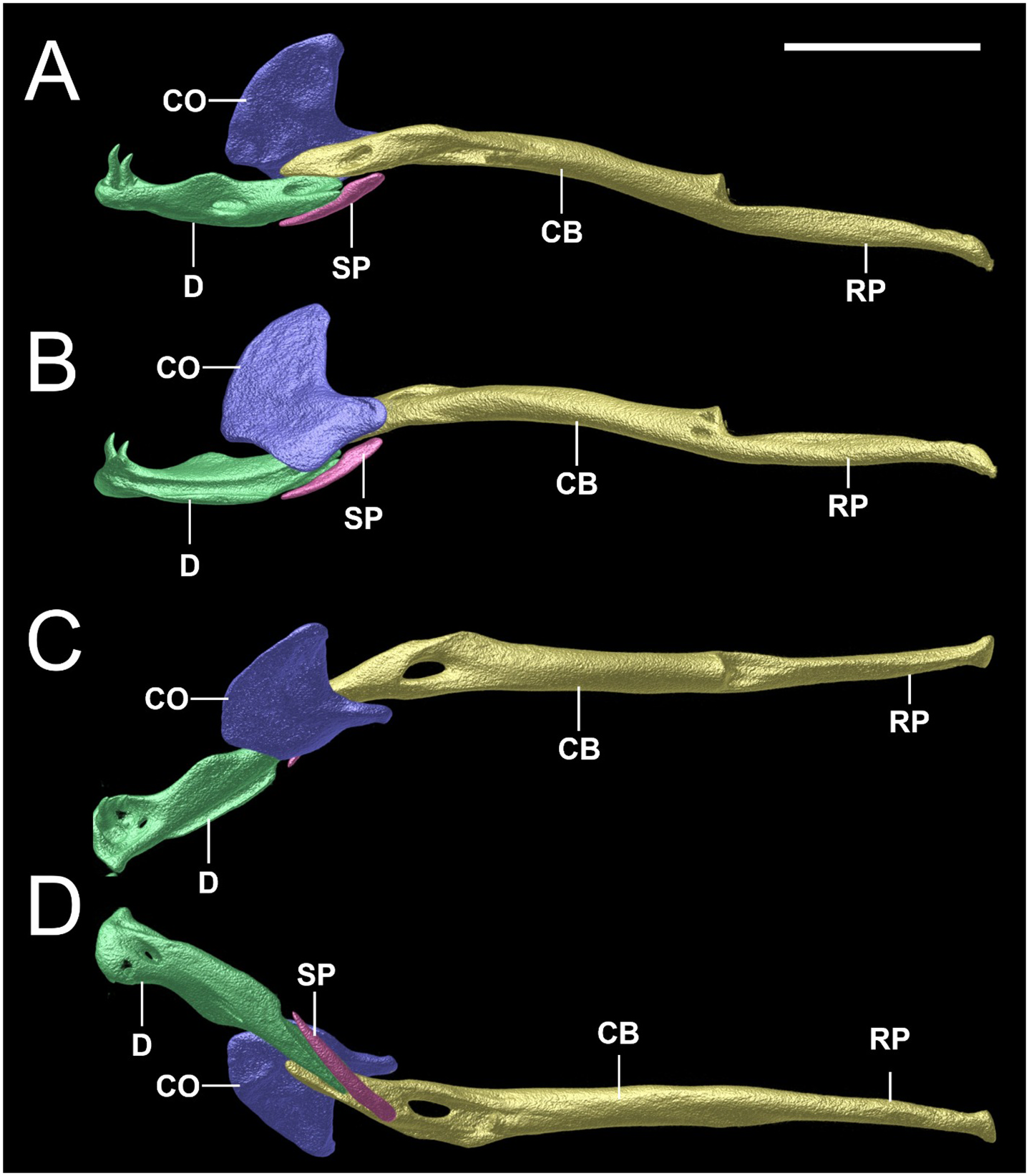
Osteological description is based upon adult paratypes of L. beui (BMNH 1946.1.11.12, MCZ-R 17842, 16702, Figs. 10–12 View Fig View Fig View Fig ). Comparisons are made to the osteological description of L. albirostris ( Rieppel et al., 2009; characters in parenthesis). In L. beui , the lateral flange of the nasal forms a more pronounced and longer projection that overlaps the laterally descending frontal flange (lateral flange of the nasals forms a less pronounced, smaller projection, slightly overlapping the laterally descending frontal flange in L. albirostris ); the foramina on the external surface of the nasal tend to be concentrated along the midline of the dorsal surface, with its anterior surface of nasal without foramina, rugose thickened (foramina on the external surface of the nasal are concentrated on both sides of the midline on its anterior and dorsal surfaces in L. albirostris ); the anterior end of nasal is pointed (straight in L. albirostris ); the anterior end of the vomer remains separate from the premaxilla, with vomers unfused along their entire length (fused on posterior part in L. albirostris ); the paired frontals meet the paired parietals in a broad V-shaped suture (U-shaped suture in L. albirostris ); the postorbital element contacts the lateral surface of the prefrontal slightly anterior to the prefrontal-frontal articulation (separated from the prefrontal in L. albirostris ); no foramina in the dentigerous process of maxilla (foramina present in dentigerous process of maxilla in L. albirostris ); the posterolateral process of palatine contacts anterior region of pterygoid (separated from pterygoid in L. albirostris ); an ossified supratemporal and supraoccipital are both absent (present in L. albirostris ); trigeminal foramen larger, formed by the parabasisphenoid, parietal and prootic (trigeminal foramen smaller in L. albirostris , formed by parietal and prootic); ventral surface of basioccipital indented at both sides of the anterolateral region (smooth in L. albirostris ); ventral margin of coronoid not in contact with splenial (identified as “angular” by Rieppel et al. (2009), contacting splenial in L. albirostris ); splenial visible in lateral and medial views, posterior surface not obscured in medial view by the compound bone and coronoid (posterior surface obscured by compound bone and coronoid in L. albirostris ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |
Liotyphlops beui
| Entiauspe-Neto, Omar M., Franco, Francisco L., Koch, Claudia, Tiutenko, Arthur, Wingert, Juliana M. & Borges-Martins, Márcio 2023 |
L. sousai
| Santos, F. J. M. & Reis, R. E. 2018: 507 |
Helmintophis beui
| Amaral, A. 1924: 25 |