
Madagascar, Montreuil, 2011
|
publication ID |
https://doi.org/ 10.5252/z2011n1a5 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC9E64-DA51-3A66-3956-FC96876EE3AD |
|
treatment provided by |
Felipe |
|
scientific name |
Madagascar |
| status |
|
«Groupe Epilissus View in CoL »
DÉFINITION TRADITIONNELLE DES GENRES EPILISSUS REICHE, 1841 , ARACHNODES WESTWOOD, 1847 , PSEUDARACHNODES LEBIS, 1953 , ET PEYRIERASANTUS PAULIAN, 1976
Le genre Epilissus a été créé par Reiche (1841) au sein de sa clé des Lamellicornes coprophages pour isoler Canthon prasinus Klug, 1833 , originaire de Madagascar . Quelques années plus tard, Westwood (1847) reprend le résultat de Reiche et introduit le nom Arachnodes pour y ranger une autre espèce malgache, A. nitidus , décrite originellement par Castelnau (1840) dans le genre Circellium Latreille, 1825 (émendé en Circelium ). Westwood ne compare pas directement les deux genres Epilissus et Arachnodes , mais donne une brève description de chacun d’eux, d’où il ressort essentiellement des différences dans la forme des pattes et du thorax pour les séparer.
Comme quelques autres noms de genre introduits par Westwood, Arachnodes sera longtemps ignoré par les auteurs ( Montreuil 2006). Il ne sera repris que bien plus tard par Olsoufieff (1947). Cependant, la distinction de ce genre avec Epilissus est peu nette dans les travaux d’Olsoufieff, la forme si particulière du pronotum mise en avant pour caractériser les Arachnodes se retrouvant également chez Epilissus
prasinus comme le souligne d’ailleurs l’auteur. En fait, il ressort des travaux d’Olsoufieff que ces deux genres ne différeraient entre eux que par la coloration, ainsi que par la structure remarquable du côté de l’élytre, formant un large pseudoépipleure chez E.prasinus . Par la suite, c’est uniquement ce dernier caractère élytral qui sera mis en avant ( Lebis 1953) pour séparer, au sein de la lignée des Canthonini brévitarses malgaches, le genre monospécifique Epilissus du genre Arachnodes , au nombre d’espèces constamment croissant.
Par ailleurs, Lebis (1953) introduit le nom Pseudarachnodes pour isoler au sein même des Arachnodes deux espèces présentant des paramères dissymétriques et bifides. Enrichi d’une troisième espèce, ce groupe, initialement défini comme sousgenre, sera élevé au rang de genre ( Paulian 1976a) et repris par la suite avec ce niveau taxinomique ( Paulian 1986; Montreuil 2003a). Deux espèces y seront ajoutées ( Montreuil 2003a).
Plus récemment, Paulian (1976c) crée le genre Peyrierasantus pour isoler P. nitidus Paulian, 1976 . Décrit originellement dans la tribu des Pinotini Kolbe, 1905 (synonyme d’Ateuchini Castelnau, 1840), ce genre sera ensuite placé par Paulian (1986) parmi les Canthonini, sans adjoindre aucune considération particulière quant à sa place au sein de cette tribu: ce taxon figure en effet à la fin de son catalogue des Canthonini malgaches, juste à la suite des genres appartenant à la lignée des longitarses, lignée à laquelle on est alors tenté de le rattacher.
MONOPHYLIE DU « GROUPE EPILISSUS »
Au sein des brévitarses malgaches, le « groupe Epilissus », composé des genres Epilissus, Archnodes , Pseudarachnodes et Peyrierasantus , pris dans leur définition traditionnelle, s’oppose au « groupe Nanos », composé des genres Nanos Westwood, 1847 , Cambefortantus Paulian, 1986 , et Apotolamprus Olsoufieff, 1947 , par leur structure élytrale simple à sept stries bien individualisées sur le disque ( Montreuil & Viljanen 2007; Montreuil 2008b) et par les côtés de leur pronotum fortement déclives près des angles postérieurs, ces derniers non visibles du dessus. D’autres Epilissini de la faune mondiale présentent des structures élytrales et une forme du pronotum similaires à celles observées chez les représentants du «groupe Epilissus », les genres d’Epilissini néo-calédoniens par exemple.
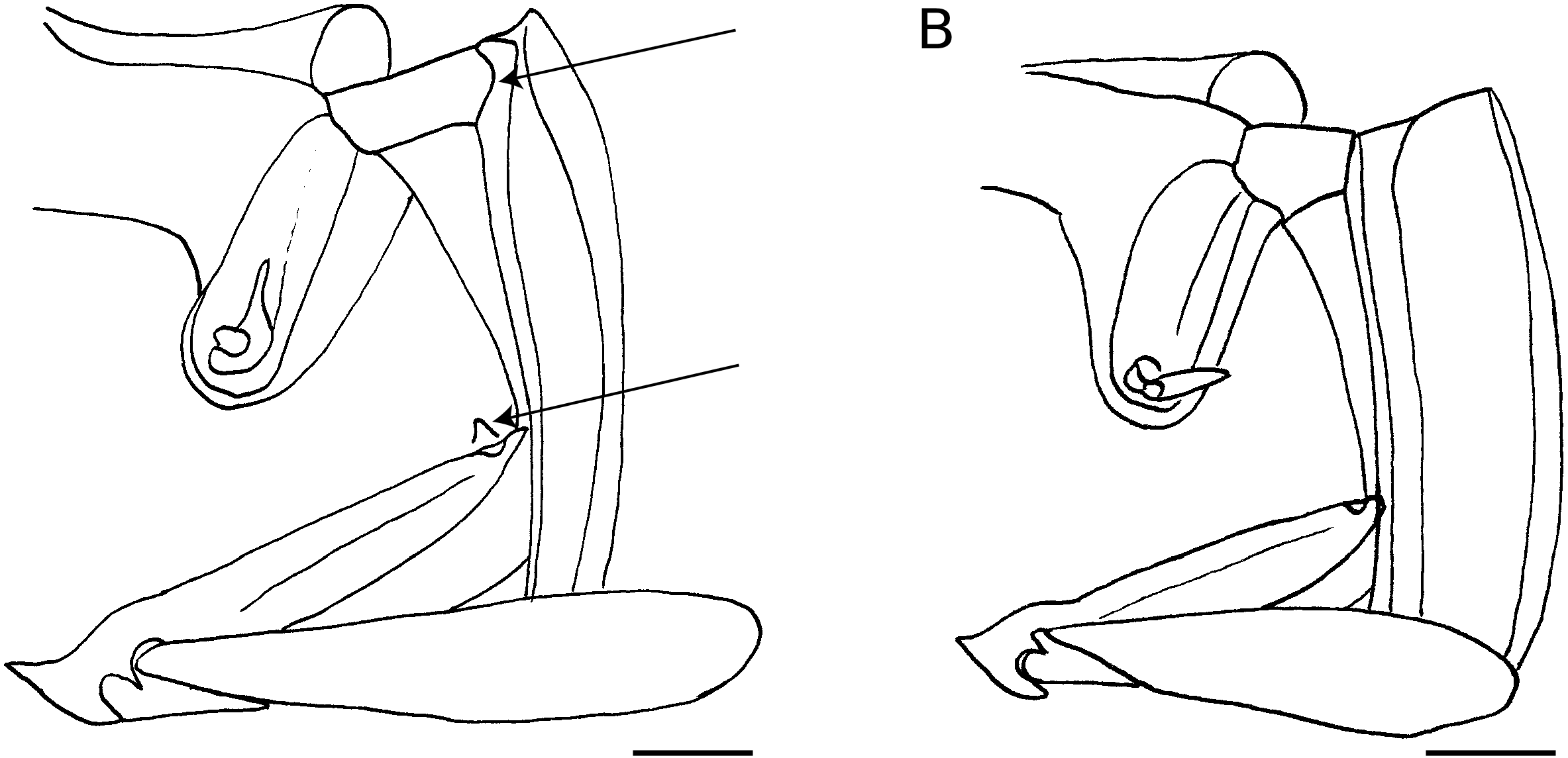
À l’exception d’une seule espèce, que nous pouvons supposer pourtant appartenir à ce groupe (voir cidessous), les espèces du «groupe Epilissus » présentent un petit relief plus ou moins saillant et caréniforme en arrière du métasternum, généralement bien visible au niveau de l’angle externe postérieur ( Fig. 1A View FIG ), structure observée, jusqu’à présent, chez aucun autre genre d’Epilissini de la faune mondiale ( Fig. 1B View FIG ). Ce caractère constituerait une apomorphie pour ce groupe et supporte l’hypothèse de monophylie de celui-ci.
Une espèce ne possède pas ce caractère, peut-être la conséquence d’une réversion. Cette espèce présente cependant des caractères communs à d’autres espèces du «groupe Epilissus », permettant de la placer avec ces dernières au sein d’un nouveau genre qui sera défini plus loin.
REFONTE DES GENRES EPILISSUS REICHGE, 1841 , ET ARACHNODES WESTWOOD, 1847 , ET MISE EN SYNONYMIE DE PSEUDARACHNODES LEBIS, 1953 Un ensemble de caractères morphologiques permet de redéfinir et de préciser les limites des genres Epilissus et Arachnodes .
Structure élytrale
La structure élytrale d’ Epilissus prasinus montre un pseudoépipleure exceptionnellement large chez cette espèce simplement du fait que le 8 e interstrie est fortement saillant, caréné. La partie externe du 8 e interstrie est ainsi projetée dans un plan vertical situé dans le prolongement du pseudoépipleure. Mise à part cette structure au niveau du 8 e interstrie, la structure élytrale d’ E. prasinus est similaire à celle observée chez tous les Arachnodes ( Montreuil 2008b) et Pseudarachnodes . Notons ici que cette forme particulière du 8 e interstrie peut être également observée, de manière plus ou moins marquée, chez d’autres genres de Canthonini malgaches, notamment chez nombre d’espèces du genre Nanos Westwood, 1847 , ainsi que chez quelques représentants du genre Apotolamprus , ce qui semble constituer autant de convergences.
A
Forme des protibias
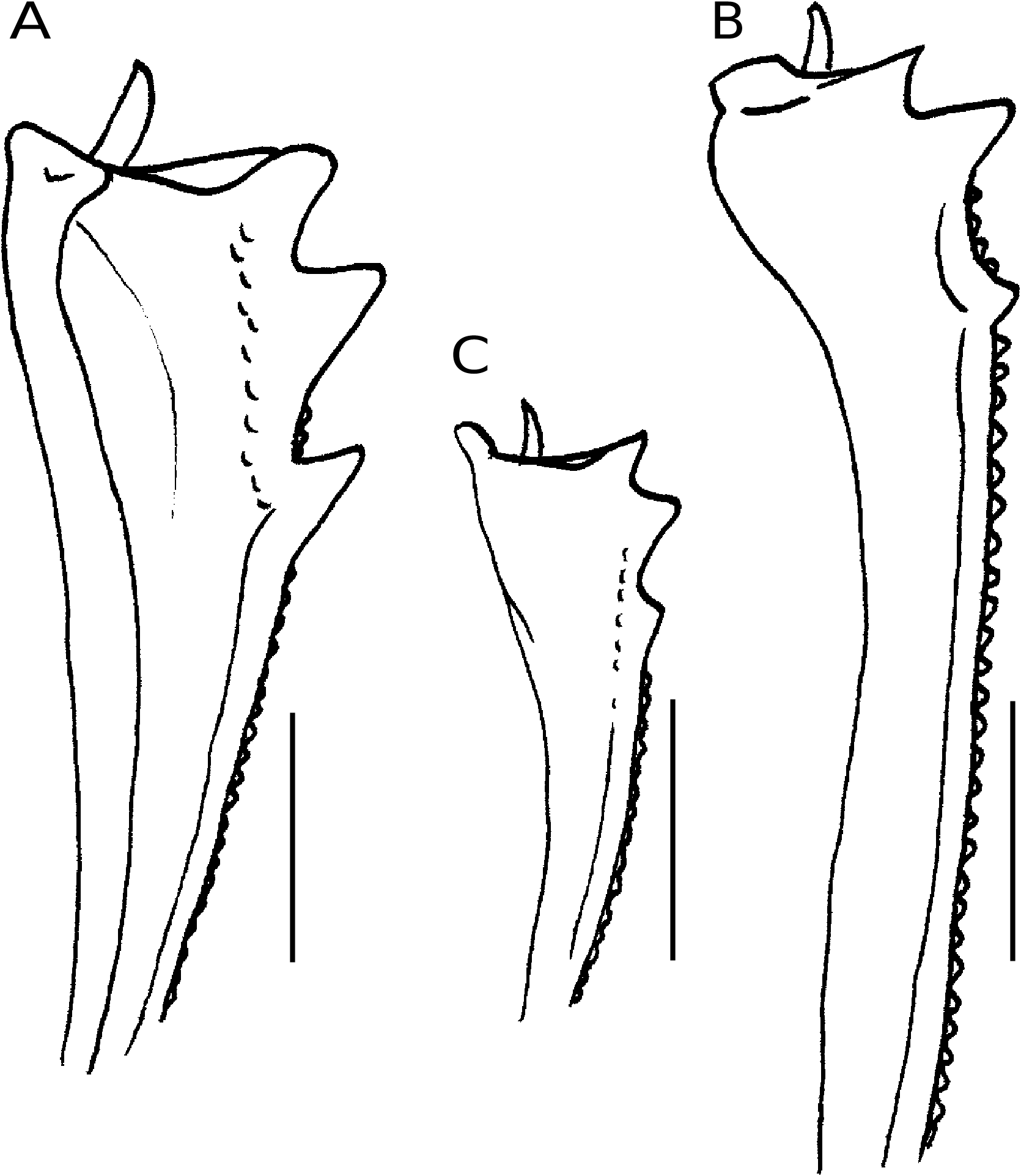
Dans sa définition des Arachnodes et Epilissus, Westwood (1847) met en avant la forme générale des protibias pour séparer les deux genres. Une partie des Arachnodes présentent des protibias certes variables de forme, mais généralement assez frêles, souvent peu élargis avant l’apex, et sans structure particulière sur la face dorsale ( Fig. 2B, C View FIG ). Sont concernées essentiellement les espèces des groupes « A. refulgens », « A. nitidus » et « A. goudoti » de Lebis (1953) , auxquelles il faut ajouter les espèces placées habituellement dans Pseudarachnodes . Ce groupe ainsi recomposé, incluant A. nitidus ( Castelnau, 1840) , espèce type du genre Arachnodes , constitue le genre Arachnodes s.s. L’autre partie des Arachnodes (groupes « A. viridis », « A. splendidus » et « A. morio » de Lebis [1953] ) présentent des protibias bien différents des précédents, assez forts, régulièrement élargis depuis la base, et ayant sur leur face dorsale un relief caréniforme longitudinal bordant la marge interne, visible pratiquement depuis la base jusqu’à l’apex ( Fig. 2A View FIG ). Cette forme des protibias se retrouve également chez Epilissus prasinus . Cette partie des Arachnodes + Epilissus prasinus constitue le genre Epilissus s.s. dans une nouvelle acception.
Forme des mésépimères
La forme des mésépimères permet également de faire la même coupe et de séparer Arachnodes s.s. + Pseudarachnodes d’ Epilissus s.s. En effet, chez les derniers, les mésépimères sont plans et leur côté externe est droit, dans le prolongement des métépisternes ( Fig. 1B View FIG ), ce que l’on retrouve habituellement chez les Epilissini et qui constituerait un état plésiomorphe pour cette tribu. Chez les Arachnodes s.s., le côté des mésépimères n’est pas dans le prolongement des métépisternes. Il est saillant par rapport au côté externe des métépisternes et plus ou moins anguleux vers l’extérieur, recouvrant légèrement une petite partie du côté de l’élytre ( Fig. 1A View FIG ). Chez certaines de ces espèces, les mésépimères sont déprimés et leur bord externe est plus ou moins relevé en bourrelet et apparaît ainsi moins saillant vers l’extérieur. C’est de ces dernières espèces que l’on doit rapprocher les espèces habituellement placées dans Pseudarachnodes , car ces dernières présentent cette structure déprimée des mésépimères avec en outre un bourrelet au bord externe plus ou moins développé. Les espèces habituellement placées dans Pseudarachnodes formeraient tout au plus un groupe naturel d’espèces à paramères disymétriques au sein d’une première lignée d’ Arachnodes s.s. à habitus massif, aux protibias normaux et aux yeux de taille normale. Cette lignée s’oppose à une deuxième lignée d’ Arachnodes s.s., présentant des protibias bien plus frêles et des yeux proportionnellement plus grands. Certaines espèces présentent cependant des caractères intermédiaires entre ces deux groupes, ce qui ne me permet pas de définir, pour le moment, une séparation claire entre ces deux lignées au sein d’ Arachnodes s.s.; c’est pourquoi je propose ici de regrouper ces deux ensembles dans Arachnodes s.s., et la synonymie entre Arachnodes Westwood, 1847 , et Pseudarachnodes Lebis, 1953 n. syn.
IMPLICATIONS TAXINOMIQUES
ET NOMENCLATURALES
Suite à la redéfinition des genres Epilissus et Arachnodes , le transfert dans le genre Epilissus d’une partie des espèces jusqu’ici placées dans Arachnodes s’avère nécessaire.Sont concernées essentiellement les espèces placées dans les trois premiers groupes d’ Arachnodes définis par Lebis (1953): groupes « A. viridis », « A. splendidus » et « A. morio ». L’ensemble de ces transferts et les nouvelles combinaisons afférentes sont présentées dans le catalogue des Epilissus , Arachnodes et Apterepilissus n. gen. à la fin de ce travail.
MISE EN SYNONYMIE DU GENRE
La position systématique du genre Peyrierasantus au sein des Epilissini malgaches n’a pas été, comme nous l’avons vu précédemment, établie de manière explicite et est demeurée fort vague jusqu’à présent. Nous pouvons observer cependant que le premier article des tarses postérieurs de la seule espèce de ce genre, P. nitidus , est plus court que le deuxième, ce qui permet de placer ce taxon parmi les Epilissini brévitarses .Sa structure élytrale et la présence du petit relief à l’angle externe postérieur du métasternum la place dans le «groupe Epilissus ». L’absence de carène sur la face supérieure des protibias la rapproche du genre Arachnodes et , plus précisément, la structure de ses mésépimères déprimés et à bord externe saillant est du type observé chez le groupe des Arachnodes incluant les Pseudarachnodes . Les caractères «particuliers» que présente cette espèce, comme la dépression frontale ou le fort sillon près de la marge interne des yeux n’ont ici qu’une valeur spécifique.
La synonymie suivante est alors établie: Arachnodes Westwood, 1847 = Peyrierasantus Paulian, 1976 , n. syn.
PLACE DES EPILISSINI BRÉVITARSES MALGACHES DANS LA FAUNE MONDIALE
L’étude de la morphologie des genres d’Epilissini brévitarses de Madagascar permet de préciser les
liens de parenté entre les différents genres au sein de la faune malgache et de regrouper ceux-ci en trois lignées principales. Ce résultat est également produit par l’étude moléculaire proposée par Wirta et al. (2010). Pour comprendre les liens de parenté entre ces lignées, et de manière plus générale entre toutes les lignées malgaches, il est nécessaire d’aborder les Epilissini dans leur ensemble, à l’échelle mondiale. En effet, les caractères permettant de définir ces lignées à Madagascar se retrouvent chacun répartis chez nombre de genres d’Epilissini de part le monde, ce qui plaide pour une origine polyphylétique de la faune malgache. Ainsi la structure élytrale type « Nanos » se retrouve également chez des genres de Nouvelle-Calédonie ( Montreuil 2008b) mais également chez les longitarses tels que les genres Epactoides Olsoufieff, 1947 , à Madagascar , et Ochicanthon Vaz de Mello, 2003 , dans la région Orientale; des genres de la région Australienne présentent des structures élytrales et des paramères comparables à ceux observés chez les Nanos malgaches. Par ailleurs, la structure élytrale de type « Epilissus »se retrouve chez des genres des régions Orientale et Australienne et certains de ces genres présentent les côtés du pronotum déclives près des angles postérieurs (cf. Temnoplectron Westwood, 1842 ; Lepanus Balthasar, 1966 ; Monoplistes Lansberge, 1874 ) observés chez le «groupe Epilissus ».
Ce travail ne constitue qu’une première approche permettant la mise en place de nouvelles hypothèses qu’il conviendra de vérifier et de tester au moyen d’études phylogénétiques plus formelles.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.