
Austrinoecia, 2011
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2857.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC87FD-EC3B-FFFA-6FDD-FF72A310F801 |
|
treatment provided by |
Felipe |
|
scientific name |
Austrinoecia |
| status |
gen. nov. |
AUSTRINOECIA Chavtur & Angel View in CoL n. gen.
Synonymy:
1906a Conchoecia rotundata group—Müller, p. 79 (part).
1908 Conchoecia —Müller, p. 77 (part).
1920 Conchoecia rotundata group—Skogsberg, p. 648 (part).
1955 Metaconchoecia —Howe, p. 118 (part).
1973 Metaconchoecia —Poulsen, p. 70 (part).
1974 Conchoecia —Deevey, p. 367 (part).
1978a Conchoecia —Deevey, p. 55 (part).
1981 Conchoecia skogsbergi species complex—Gooday, p. 159 (part).
1982a Conchoecia rotundata group—Deevey, p. 148 (part).
1982 Conchoecia —Jazdewski et al. (part).
1986 Metaconchoecia —Kempf, p. 486 (part).
1992 Metaconchoecia —Kock, p. 68 (part).
1992 Conchoecia —Benassi et al. p. 71 (part).
1997 Metaconchoecia —Blachowiak-Samolyk and Zmijlewska, p.80 (part).
1997 Metaconchoecia —Gollasch, p. 224 (part).
1999 Metaconchoecia —Angel, Figs 9.58 View FIGURE 9 –76 (part).
2002 Metaconchoecia —Blachowiak-Samolyk and Osowiecki, p.140 (part).
2003 Metaconchoecia —Chavtur and Kruk, p. 170 (part).
2007 Metaconchoecia —Blachowiak-Samolyk and Angel p. 71 (part).
Etymology. The name is derived from the Latin adjective austrinus, meaning southern, and “-oecia” derived from the Greek word “οƖκοσ” meaning house, from which the terms ecology and economy have been derived, and is the ending that has become standard for Conchoeciinae genera.
Type species. Conchoecia isocheira ( Müller, 1906a) View in CoL .
Composition. The genus is monospecific, containing only Austrinoecia isocheira ( Müller, 1906a) View in CoL a species that is endemic to the Southern Ocean.
Differential diagnosis. Small (0.76–1.12 mm), deep mesopelagic species. Females tend to be slightly larger than males. The maximum carapace height is just anterior to midlength in males, but is just posterior to midlength in females. The LAG opens just posterior to the anterior end of the hinge. The RAG opens at the apex of an angle in the posterior margin at about 15% CH below the hingeline. The d-seta on the male A1 is longer than the e-seta. The e-seta armature consists uniquely of a single row of 7–9 hyaline flanged spines. The copulatory appendage has a characteristic deep terminal indentation on its anterior margin.
Description. Males. Carapace. The length range is 0.76–0.95 mm (mean 0.86 ± 0.036 mm). The carapace is relatively short and high. The maximum height of 50–61%CL occurs anterior to midlength. The breadth can exceed the height in some specimens, but it is generally 53–54%CL. The LAG opens 12–15%CL behind the tip of the rostrum and 3.5%CL posterior to the anterior end of the hinge. The RAG opens on the apex of a slight protuberance on the posterior margin 13–17% CH below the dorsal hinge line. The rostrum is quite well developed, 7– 9%CL. There is no surface ornamentation.
Frontal organ: There is a clear septum dividing the capitulum from the shaft. The shaft is quite short (27%CL), and the capitulum is even shorter (12%CL). It is barely thicker than the shaft. It is either straight or slightly concave, and its tip is rounded. The end of the capitulum barely extends beyond the end of the A1.
First antenna. The limb is clearly segmented and is about 3x the length of the capitulum. The a-seta is simple and extends back almost to the proximal end of the second segment. The c-seta is similar in length or slightly longer than the total combined length of the third, fourth and fifth segments (5.5%CL). The e-seta is similar in length (36%CL) to the limb. Its armature is unique in that it consists of a single row of 7–9 hyaline, flanged spines, in contrast to the double row seen in all other Metaconchoeciini species. The length of the d-seta is also exceptional for the tribe in being longer (1.2x) than the e-seta.
Second antenna. The protopodite is 46%CL. The first segment of the exopodite is relatively slim, about 35% the length of the protopodite. The h–j sensory setae on the endopodite are 50–60% of the length of the g-seta. The g-seta is 50–62% of the length of the protopodite, and 73–74% of the exopodite, and 1.7–1.8x the first endopodite segment. Both the f- and g-setae are more or less parallel-sided with either rounded or pointed tips. The h-seta is simple and undivided. The c- and d-setae are stout and quite long, but the e-seta is minute. The hook appendages on the endopodites are curved and terminally rounded. They are similar in size and shape, with large, stout distal arms that are swollen centrally.
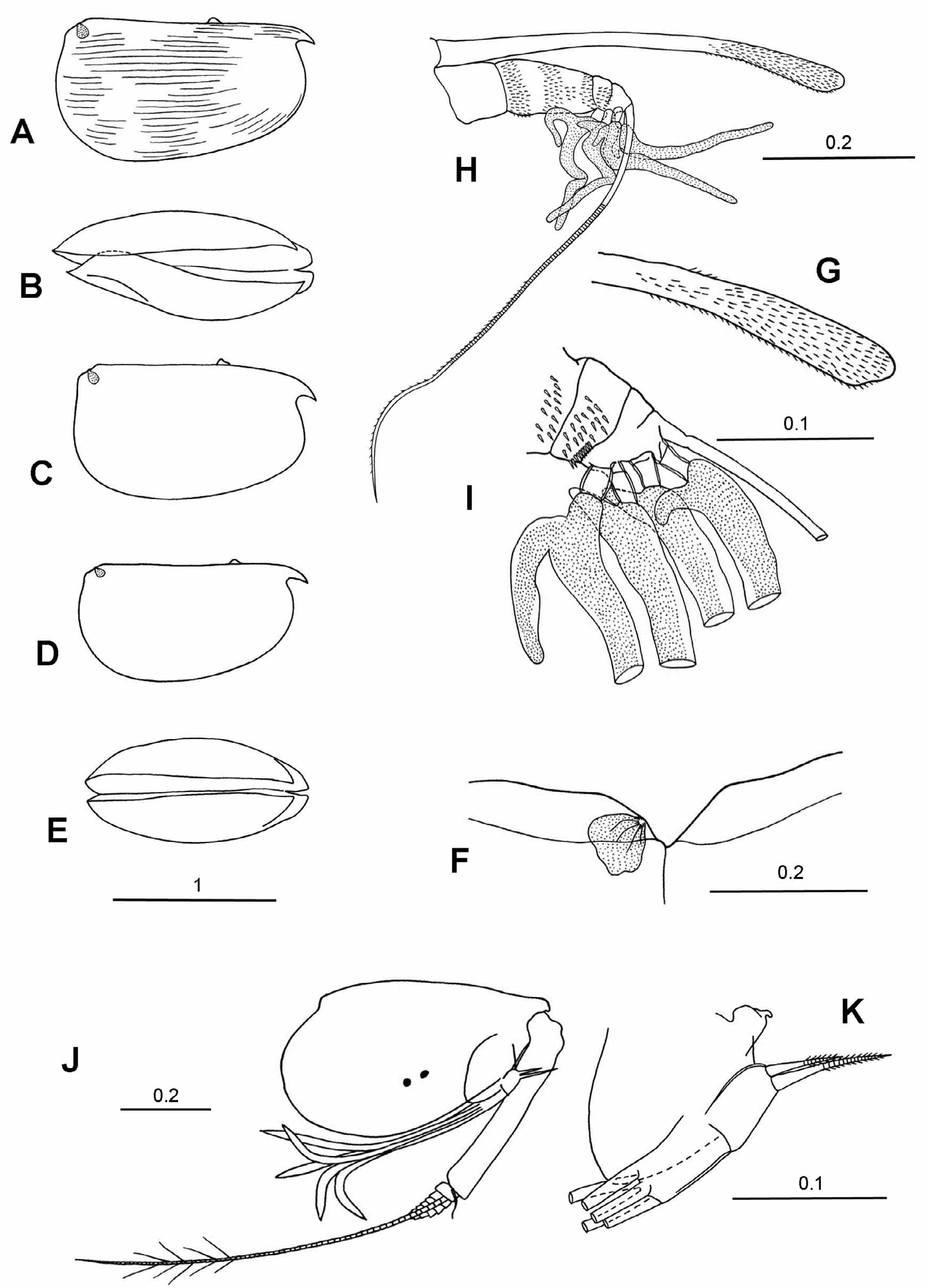
Mandible ( Fig. 2L View FIGURE 2 ). The exopodite is reduced to a minute triangular protuberance that carries a long, slim, seta with long setules. The coxale is relatively short and about two-thirds the length of the endopodite. The first segment of the endopodite has a finely spinose, subterminal seta on its dorsal surface and two setae on its inner ventral surface, one longer than the endopodite, the other half its length. The longest terminal seta of the endopodite is 22%CL and nearly 90% of the endopodite itself.
Copulatory appendage. The appendage is large, being 45–48%CL. It is highly curved distally and tapers terminally. The tip characteristically carries an offset triangular plate. The appendage is broadest near or slightly beyond midlength. There are 2–4 oblique muscle bands.
Females. Carapace. The length range is 0.90–1.12 mm (mean 1.01 ± 0.041 mm). The carapace is short and more elongate than in the male (42%CL); the maximum height occurs at or just behind midlength. The breadth is 48%CL. The LAG opens 12–15%CL behind the tip of the rostrum and 3%CL posterior to the anterior edge of the hinge. The RAG opens on a small protuberance on the posterior margin 12–17% CH below the posterior dorsal corner. The rostrum is less developed than in the male, 5–8%CL. As in the male, there is no sculpture on the carapace. Frontal organ. The shaft is clearly divided from the capitulum by a septum. The shaft is 18%CL. The capitulum is 12%CL; it is straight and narrow with a rounded or bluntly pointed tip and is sparsely spinous around the flanks, especially at midlength. Its thickness is similar to or slightly larger than that of the shaft. The whole frontal organ is 29–30%CL, and is 1.5x the length of the A1.
First antenna. The limb segments are fused, and their total length is 20–21%CL. There is no dorsal seta. The aseta is simple and undivided. The e-seta is 1–1.5x the length of the limb and 1.3–1.8x the sensory setae. Distal to the ends of the sensory setae, its trailing edge is armed with strong, long spines.
Second antenna. The protopodite is 42–43%CL. The first exopodite segment is 20%CL and relatively thin; segments 2–9 that carry the swimming setae are 44% of the first segment. The terminal setae on the endopodite are thin, and disparate in length, (f-seta 20%CL, g-seta 22.5%CL, and the h- to j-setae 19%CL). They are either parallel-sided or very slightly tapered, with pointed tips.
Mandible. The structure of this limb is similar to that of the male.
Distribution. Endemic to the Southern Ocean where it occurs over a wide latitudinal range from 49ºS to 78ºS in all sectors ( Müller 1906a, 1906c, 1908; Skogsberg 1920; Hillman 1967; Deevey 1974, 1978a, 1982a, 1983; Angel 1981b, Hopkins 1985; Hopkins & Torres 1989; Kock 1992; Naldi et al. 1992; Benassi et al. 1992; Blachowiak-Samolyk 1999, 2001; Blachowiak-Samolyk & Angel 2003, 2007: Blachowiak-Samolyk & Zmijewska 1995, 1997; Blachowiak-Samolyk & Osowiecki 2002; Drapun 2004). Its overall depth range is reported to be 0–3248m. However, the deeper records are all from nets fished open. Stratified sampling in the austral summer has shown it generally occurs between 200–500m and occasionally as shallow as 100m. The possibility that it undertakes seasonal migration into deep water in winter can not be ruled out.
CLAUSOECIA Chavtur & Angel n.gen.
Synonymy
1906a Conchoecia ‘rotundata’ group—Müller, p. 79 (part).
1955 Metaconchoecia —Howe, p. 118 (part).
1968 Conchoecia rotundata group—Deevey, p. 50 (part).
1973 Metaconchoecia —Poulsen, p. 70 (part).
1980 Conchoecia rotundata group—Deevey & Brooks, p. 85 (part).
1981 Conchoecia skogsbergi species complex—Gooday, p.141 (part), 159 (part).
1982a Conchoecia rotundata group—Deevey, p.148 (part).
1986 Metaconchoecia —Kempf, p. 486 (part).
1992 Metaconchoecia —Kock, p 68 (part).
1993 Conchoecia —Angel, p. 204 (part).
1999 Metaconchoecia —Angel, Figs 9.58 View FIGURE 9 –76 (part).
Etymology. The name is derived from “Claus” in honour of that early doyen of halocyprid taxonomy Carl Claus, and “-oecia” derived from the Greek word “οƖκοσ” meaning house, from which the terms ecology and economy have been derived, and is the ending that has become standard for Conchoeciinae genera.
Type species. Conchoecia pusilla ( Müller, 1906a) View in CoL .
Composition. The genus is currently monospecific containing just C. pusilla ( Müller, 1906a) View in CoL but significant size differences have been reported between different populations. Müller 1906a, for example, reported a small form, so probably there are cryptic species awaiting recognition.
Differential diagnosis. Small, deep mesopelagic species <1 mm in length with the maximum height posterior to midlength. The LAG opens 12 to 15%CL posterior to the tip of the rostrum. The RAG opens on the apex of an angle in the posterior margin 10 to 12%CL behind the posterior margin of the hinge. The carapace has a slight anterior taper and the flanks curve smoothly. The male A1 e-seta armature consists of 20 to 25 slim spines that are paired distally and become alternate proximally.
Description. Males. Carapace: The length range is 0.66–1.00 mm. Müller (1906a) in his original description, described the males as being typically 0.85–0.95 mm in length, but also reported a small ‘form’ in which the males are 0.75–0.8 mm. These smaller forms are yet to be investigated. The carapace is somewhat elongate, with a maximum height of 45–50%CL, located close to midlength. The dorsal margin is nearly straight, and the ventral margin is slightly concave, so that the anterior and posterior halves of the carapace are very similar. The maximum breadth of 47%CL is slightly anterior to midlength, and in many specimens it exceeds the height. Lrost is 9.3%CL, but LC3 is 7%CL. The LAG opens 14–15%CL posterior to the tip of the rostrum, and nearly 6%CL behind the posterior margin of the incisure. The RAG opens at the apex of a distinct angle in the posterior margin 12.5%CL behind to the posterior end of the hinge and 11–15% CH. The posterior margin dorsal to the RAG is straight and forms an obtuse angle with the dorsal margin at the posterior margin of the hinge. There is no surface ornamentation.
Frontal organ: The shaft is similar in length (33.4%CL) to the A1 limb, and is separated from the long (20.1%CL) capitulum by a suture. The capitulum is long, narrow, down-turned and bare. The basal two-thirds of its length is almost straight-sided, but distally the dorsal margin is convex, so the capitulum broadens towards its rounded end.
First antenna: The limb is segmented, and the first segment is slightly smaller than the second. The dorsal seta that locks the shaft of the frontal organ to the second segment of the limb is located 14.8%CL from the base of the capitulum. The a-seta is simple, undivided and quite long (27.8%CL). It extends well beyond the suture between the first two segments. It is S-shaped but unswollen at its base. The c-seta is short (6.2%CL), and shorter than the combined length of the third, fourth and fifth segments. The e-seta is 39%CL, only slightly longer than the b-seta (38%CL) and the d-seta (35.5%CL). The e-seta armature consists of 20–25 broad-based, slightly curved, pointed spines, which are paired distally, but become alternate proximally.
Second antenna: The protopodite is relatively long (about 58%CL). The first exopodite segment is 20%CL and has a terminal seta that curls around the base of the second segment. Together, segments 2–9 are 45% the length of the first segment, so the whole exopodite is 60–65% the length of the protopodite. On the endopodite, the g-seta is 43–44%CL, and the f-seta is 36%CL; both these setae are slightly flattened terminally and have pointed ends. The sensory h-, i-, and j-setae are simple, thin-walled and similar in length (14–15%CL). The hook appendages are slightly curved and end in oblique points with subterminal ridging. The left hook is slightly smaller and straighter.
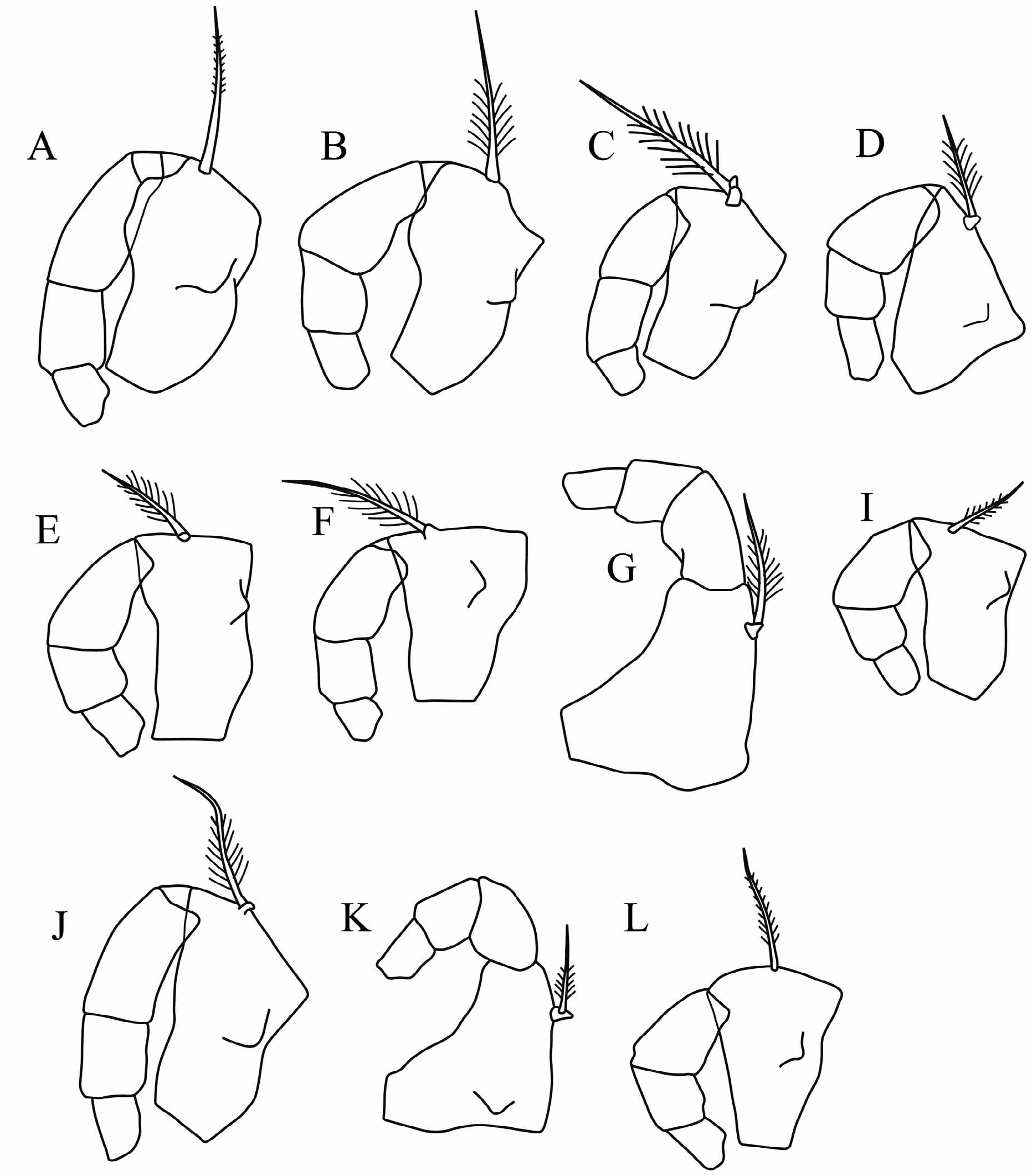
Mandible ( Fig. 2E View FIGURE 2 ): Dorsally the first endopodite segment has a subterminal, short, finely seta with long setules. On its inner ventral face this segment has a single, moderately long seta that does not quite reach to the end of the limb. As usual there are seven terminal seta on the third segment; the longest is 22%CL and similar in length to the limb itself. The exopodite consists of a short process armed with short seta with long setules, which is quite stout at its base but rapidly becomes quite slim. The coxale is short and about half the length of the endopodite.
Maxilla: The setation of the endopodite is consistent with all the other species in the tribe. On the basal segment there are four anterior setae, a single lateral seta and three posterior setae. The terminal segment carries three hook setae and two lateral setae.
Fifth limb: The epipodial appendage has three groups of 4 + 4 + 4 setae. The basale generally has ventrally a group of 5 basal setae and a further 2 setae distally. This segment has no lateral setae, but occasional specimens do appear to have three lateral setae but correspondingly fewer ventral setae. The orientation of the limb on the slide probably determines whether these setae appear to be lateral or ventral. Dorsally, there is a long subterminal seta, but no seta with setules was seen in the specimens dissected. Drapun (personal communication) has observed a dorsal seta with long setules. The first endopodite segment has two medial ventral setae and one medial dorsal seta. All three of these setae are quite long and extend well beyond the end of the segment. The terminal setae have length ratios of 100:95:50; it is unusual for the dorsal terminal seta to be the longest. The longest seta is 10.2%CL and 70% of the limb.
Sixth limb: The setae on the epipodial appendage are as usual arranged in three groups of?6 + 5 + 5. The basale has 3 or 4 ventral setae, 0 to 2 lateral setae and dorsally a single seta with long setules. The first endopodite segment has a single short, medial seta ventrally but no dorsal seta. The second segment has a single short medial seta on both the dorsal and the ventral sides. The three long terminal setae are similar in length (about 38%CL), and>120% the length of the limb.
Caudal furca: There are the usual eight pairs of spines on the caudal furca, the longest of which is 15.5%CL. There is an unpaired seta posterior to the paired spines.
Copulatory appendage: The length of the appendage is 34%CL. The posterior edge is almost straight, and the ventral edge is slightly convex in its distal half. The end is bluntly rounded, and there are four or five oblique muscles.
Females. Carapace: The length range is 0.7–1.0 mm. The carapace is elongate and tapers very slightly, although the maximum height of 45–53%CL is located close to midlength. In ventral aspect the carapace is slim and spindle-shaped; its maximum width of 40%CL length is anterior to midlength, and the flanks are smoothly curved. The rostrum is well developed (Lrost =11%CL, LC3 = 6%CL). As in the male the LAG opens 11–16%CL posterior to the tip of the rostrum. The RAG opens at a well-marked angle in the posterior margin 10%CL behind the posterior end of the hinge and 6–8%CL below the hinge-line. Above the gland opening the posterior margin is straight forming an obtuse angle with the dorsal edge; below the RAG opening it curves smoothly into the ventral margin. The carapace has no sculpture.
Frontal organ: The frontal organ is fused and its overall length is 30%CL, and is nearly double the length of the A1 limb. The capitulum is long, straight, ventrally hirsute, and very slightly expanded distally. It has a bluntly rounded tip.
First antenna: The limb is fused with no obvious segmentation, 15%CL in length. There is no dorsal seta. The sensory a-, b-, c- and d-setae are simple and thin-walled, and are 12%CL in length. The e-seta is about double the length of the sensory setae (24.5%CL) and 1.6x the length of the limb. Distal to the sensory seta, the e-seta is lined with short fine spines along its trailing edge and also has a few longer, thicker spines along its leading edge.
Second antenna: The protopodite is 43%CL, and the first exopodite segment is 20–21%CL and relatively slim. The distal segments of the exopodite that carry the swimming setae together are about 43% the length of the first segment, making the overall length of the exopodite 65–70% of the protopodite. The terminal setae on the endopodite are simple and unflattened. The g-seta is 20%CL and the other four setae are 18%CL.
Mandible: Similar to that of the male.
Fifth limb: The epipodial appendage carries three groups of 4 setae. The basale has 3 + 2 ventral setae, two lateral setae, and dorsally a further two setae, one long and bare and the other shorter with long setules. The first endopodite segment has a pair of medial ventral setae, one extending to the end of the segment, the other a little further. There is also a single medial dorsal seta, which extends beyond the end of the segment. The length ratios of terminal seta on the second segment are 100:95:55. So as in the male, the dorsal terminal seta is the longest of the three, and is 8.8%CL and 60% the length of the limb.
Sixth limb: There are the usual three groups of epipodial setae (6 + 5 + 5 ventral). Ventrally the basale has 2 bare and 2 setae with setules. There is no lateral seta, but there is a single, long, dorsal seta with setules (Drapun observes two dorsal setae with long setules in specimens from the Indian Ocean, pers.comm.). The first segment has a single medial ventral seta. Medially the second segment has both a dorsal seta and a ventral seta; both are quite long and extend beyond the end of the segment. The length ratios of the three terminal claw setae are 80:100:65. The longest central seta is nearly 11%CL and 56% of the limb.
Caudal furca: There are eight pairs of claw setae and an unpaired, slim seta at the dorsal margin of the two lamellae. The longest claw seta is 14–15%CL.
Comments. The original description of Conchoecia pusilla ( Müller, 1906a) was based on material collected from both the Atlantic and the Indian Oceans, in which Müller reported collecting a small form var. minor from near the equator in the Indian Ocean. Our material collected from near the equator (0°, 23°W) in the Atlantic is similar in size to his small form, whereas the material collected from the vicinity of the Azores Front (35°N, 30°W) was similar in size to Müller’s typical form. Another, even smaller form has been collected from depths> 2000m near Bermuda (32°N 65°W). Another collection from the Celebes Sea contained specimens that are clearly related to C. pusilla , but look different from the typical Atlantic forms. When the taxonomy of these various forms is sorted out, the genus will probably consist of several species.
Distribution. Atlantic Ocean: between 60ºN and 3ºS ( Müller 1906a; Angel & Fasham 1975; Angel 1977, 1979, 1981b; Deevey 1968; 1978 b, 1982a; Ellis 1985) and in the area 34º–36ºS and 17º–18ºE ( Müller 1906a). Indian Ocean: from 9ºN to 23ºS ( Müller 1906a; Leveau 1967; Hanai et al. 1980). Pacific Ocean: in the area bounded by 28–29ºS and 177º–179ºE ( Poulsen 1973), in the China Seas ( Chen et al. 1983; Chen & Lin 1994, 1995), and to south of 36ºS ( Deevey 1983). Southern Ocean: recorded from 64ºS, 85ºE in the Indian Ocean sector ( Müller 1908) and from 44ºS, 178°W and 45ºS, 160°E in the Pacific sector ( Deevey 1982a). The overall reported depth range is 0–2500 m, but depth stratified samples collected in the North Atlantic show the species is mostly restricted to depths of 700–1500m.
Comparisons. Species attributed to Clausoecia and Austrinoecia are very similar in their external appearances. The main characters that discriminate these two genera are summarized in Table 7 and Appendices 1–3, and are in part illustrated in Figure 2 View FIGURE 2 .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.