
Rotundoecia, 2011
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2857.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5293187 |
|
persistent identifier |
https://treatment.plazi.org/id/03DC87FD-EC22-FFE2-6FDC-FBB8A2B6FE22 |
|
treatment provided by |
Felipe |
|
scientific name |
Rotundoecia |
| status |
gen. nov. |
ROTUNDOECIA Chavtur & Angel View in CoL , n.gen.
Synonymy:
1894 Conchoecia rotundata —Müller, p. 229 (part).
1906a Conchoecia rotundata group—Müller, p. 83 (part).
1909 Conchoecia rotundata group—Fowler, (part).
1953 Conchoecia —Iles, p. 271 (part).
1968 Conchoecia —Deevey, p. 55 (part).
1973 Metaconchoecia —Poulsen, p. 74 (part).
1974 Conchoecia —Deevey, p. 364 (part).
1978 Conchoecia —Deevey, p. 53 (part).
1979 Metaconchoecia —Martens, p. 352 (part).
1981 Conchoecia skogsbergi species complex—Gooday, p. 141 (part).
1981 Conchoecia —Angel, p. 560 (part).
1986 Metaconchoecia —Kempf, p. 486 (part).
1999 Metaconchoecia —Angel, figs 9.58–76 (part).
Etymology. The name is derived from the Latin adjective “ rotundus, -a, -um” meaning round, and “-oecia” derived from the Greek word “οƖκοσ” meaning “house”, from which the terms ecology and economy have been derived, and is the ending that has become standard for Conchoeciinae genera.
Type species. Conchoecia teretivalvata Iles, 1953 View in CoL .
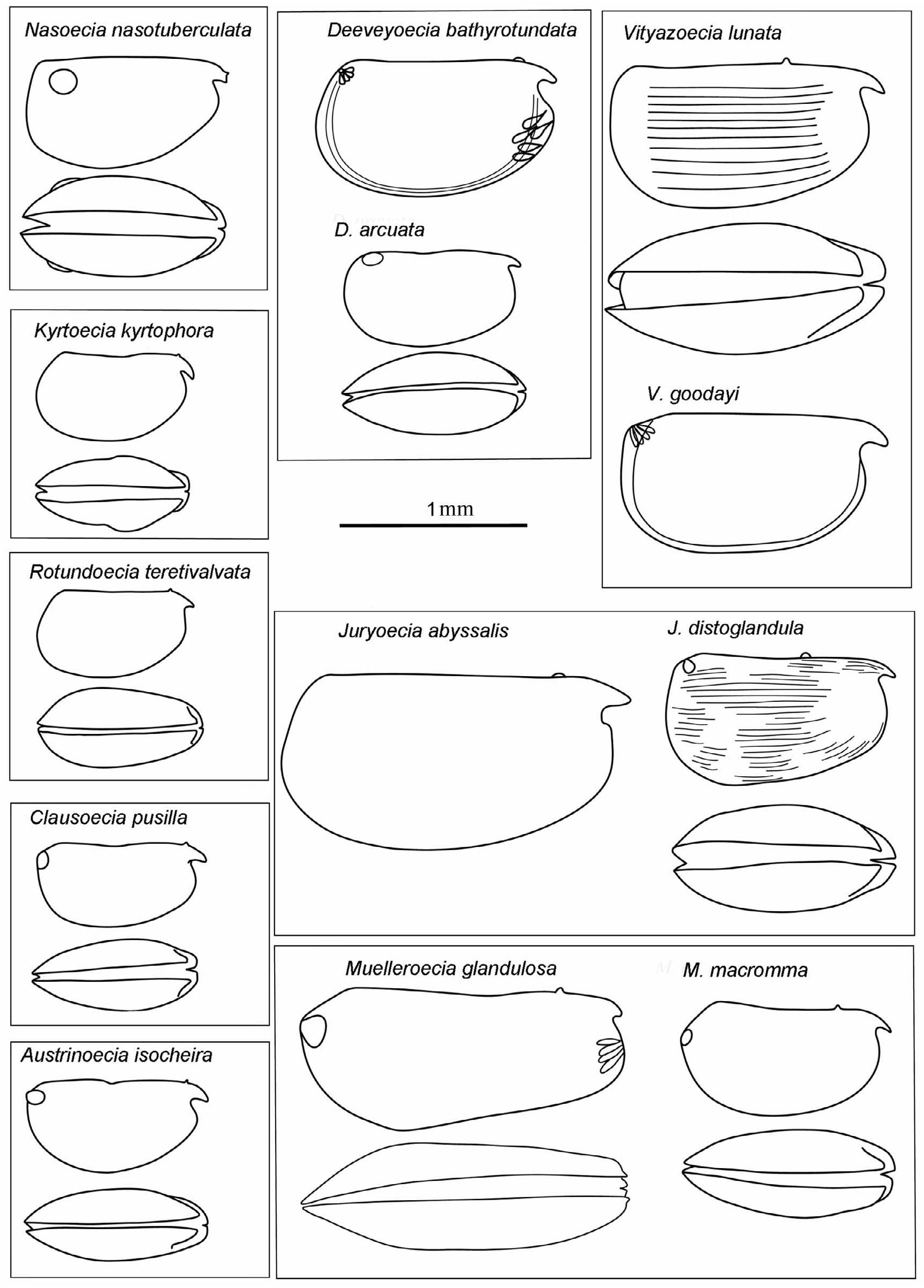
Composition. The genus consists of a single species Rotundoecia teretivalvata ( Iles, 1953) View in CoL
Differential diagnosis. Small globose species 0.78–1.15 mm, with a height of 52–60%CL and a width of 42– 48%CL. The ventral margin curves smoothly into the posterior margin, and the flanks also curve smoothly from the rostrum to the posterior margin. The carapace lacks sculpturing. The rostrum is quite small (8%CL) and the incisure is quite shallow. The LAG opens 11 to 12%CL behind the tip of the rostrum. The RAG opens on the posterior margin, nearly adjacent to the posterior end of the hinge. The male A1 e-seta armature consists of 9 to 11 pairs of slim spines.
Description. Males. Carapace: The length range in the Atlantic is 0.76–0.92 mm, but larger specimens (up to 1.15 mm) have been reported both in the Southern Ocean ( Deevey 1974) and in the S.E. Pacific off Chile ( Martens 1979). The maximum height, 50–55%CL is located well posterior to midlength, so the carapace tapers anteriorly. In the ventral aspect, the flanks curve smoothly with the maximum breadth of 48%CL at midlength. Lrost is 8%CL anterior, and LC3 is 7%CL. Ventrally the incisure is quite shallow, and the anterior margin of the carapace hardly projects beyond the inner margin of the incisure. The LAG opens 10–13%CL behind the tip of the rostrum, and 3%CL behind the anterior end of the hinge; it is also posterior to the inner margin of the incisure. The RAG opens almost adjacent to the posterior end of the hinge and close to the posterior dorsal corner. There is no visible surface ornamentation.
Frontal organ: There is a clear division between shaft and capitulum. The shaft is 32%CL, and slightly shorter than the A1 limb; it is segmented at midlength just posterior to where the locking dorsal seta from the A1 entwines around the shaft. The spinose capitulum (12.5%CL) is slightly down-turned, and similar in shape to those of Metaconchoecia species. The overall length of the frontal organ is 43–44%CL, so the tip of the capitulum just extends beyond the end of the A1 limb.
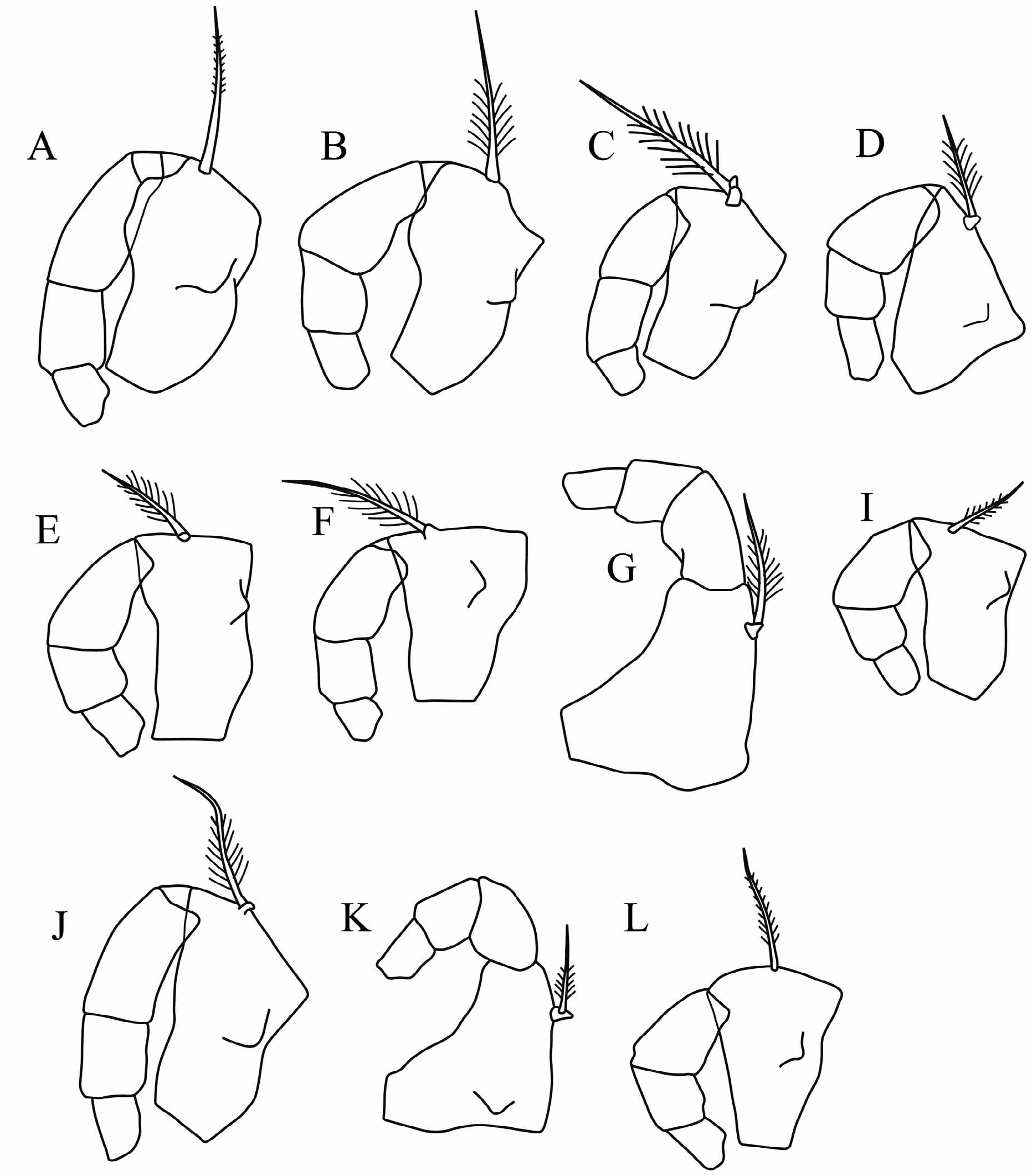
First antenna: The limb is clearly segmented, with the first segment being three-quarters the length of the second. Its overall length is 37–38%CL. The a-seta is sinuate and slightly swollen near its base. It is long (26%CL), reaching well beyond the suture between the first and second A1 segments. The c-seta is very short (4%CL). The eseta is 55%CL and is longer than both the b-seta (48%CL) and the d-seta (50–51%CL). Its armature consists of 9– 11 pairs of long, thin spines, which distally are closely spaced distally and become more spaced out proximally.
Second antenna: The protopodite is relatively long (53%CL). The slim first exopodite segment is quite short (19–20%CL) and appears to lack a terminal seta. The remaining exopodite segments are just over 50% the length of the first segment. The longest of the swimming setae is slightly longer than the protopodite. On the endopodite the g-seta is 45%CL and the f-seta is 38%CL; both are flattened and pointed. The c- and d-setae are slim and short, but the e-seta is moderately long. The h-seta is simple and subequal to the other sensory setae, i.e. 17%CL. Both hook appendages are angled and terminally rounded; the distal arm of the smaller left hook is strongly curved.
Mandible ( Fig. 2D View FIGURE 2 ): The first endopodite segment has a single subterminal dorsal seta with fine setules. It also has a ventral seta, which extends almost to the end of the limb. The longest claw seta on the end of the third segment is>90% the length of the limb and 22–23%CL. The exopodite is a well developed process, which bears a slim, medium-length seta with long setules. The coxale is relatively short and about two-thirds as long as the endopodite.
Maxilla: The endopodite has the usual setation for the tribe: four anterior setae, a single lateral seta, and three posterior setae. Terminally on its inner edge it has five small, sharp spines. The second segment has three terminal claw setae and two lateral slim setae.
Fifth limb: The epipodial setae are in three groups of 4 setae. The basale has 3+1 ventral setae, 2+2 divergent lateral setae, and dorsally both a seta with long setules and a longer, bare, subterminal seta. On the first endopodite segment there is a pair of ventral medial setae, and a single medial dorsal seta. The terminal claw setae on the second segment have length ratios of 70:100:50 (dorsal to ventral). The longest of these three setae is 8%CL and 65% the length of the limb.
Sixth limb: The epipodial setae are in three groups of 6+5+5. There are four ventral setae (1+1+2) on the basale, a single lateral seta, but no dorsal seta. The first endopodite segment has only a small medio-ventral seta. The second endopodite segment has a short dorsal and a short ventral seta; both are inserted at two-thirds of the length. The three subequal terminal setae are 47%CL and about 130% the length of the limb. All three have long setules distally.
Caudal furca: The longest pair of the eight pairs of claw setae is nearly 17%CL. Dorsally there is an unpaired seta, which is nearly quarter of the length of the longest claw setae.
Copulatory appendage: The length of the appendage is 28%CL. It is very broad, distally rounded, and terminally strongly tapered. There is a distodorsal stylet near its tip. It has 7 or 8 oblique muscle bands.
Females. Carapace: In the Atlantic the length range is 0.76–0.86 mm; larger specimens of up to 1.20 mm reported from the Southern Ocean ( Deevey 1982a), may be a second species. The carapace is short, relatively high and slightly elongated. The maximum height of 52–60%CL is located posterior to midlength, so the carapace tapers anteriorly. In ventral aspect the flanks curve smoothly. Uniquely the maximum width (42–47%CL) is located at about two-thirds of the length of the carapace. Lrost is 8 to 9 %CL, but LC3 is only about 4%CL. The anterior margin of the carapace below the incisure barely arches anterior to the inner face of the incisure. The LAG opens about 11%CL behind the tip of the rostrum, and 2–2.5%CL behind the anterior end of the hinge. The RAG opens immediately below the posterior end of the hinge near the posterior dorsal corner. There is no surface sculpture.
Frontal organ. The shaft and capitulum are fused, so the capitulum is poorly differentiated, although it is a little broader than the shaft, and is slightly down-turned. The capitulum is finely hirsute along its ventral margin, and its end is blunt or slightly pointed. The overall length of the frontal organ is 37%CL and over double the length of the A1 limb.
First antenna. The length of the limb is 17%CL, and the basal segments are fused. There is no dorsal seta. The e-seta is 35%CL, similar in length to the frontal organ. Distal to the ends of the sensory setae, the trailing edge of the e-seta is lined with fine spines. The sensory a-, b-, c- and d-setae (19%CL) are thin-walled and simple.
Second antenna. The protopodite is 49%CL. The first exopodite segment is slim and short (20–21%CL). Unlike in the male, the female appears to lack a terminal seta on the first segment. The combined length of the remaining segments of the exopodite that carry the swimming setae is about half the length of the first segment. The longest swimming seta is a little shorter than the protopodite (44–45%CL). The terminal setae on the endopodite are subequal in length (20–21%CL), thin, and parallel-sided, with rounded tips.
Mandible and Maxilla. Same as in the male.
Fifth limb. The setal formula of the epipodial appendage is the same as in the male. The basale has five ventral setae (4 +1), no (?) lateral seta, and the usual two dorsal setae, one with long setules and the other bare and longer. The first endopodite segment has a pair of medial ventral setae and a single medial dorsal seta. The length ratios of the terminal seta of the second segment are 78:100:76; the longest seta is 7.6%CL and 65% the length of the limb.
Sixth limb. The epipodial appendage has three groups of setae, 6+5+5 as in the male. The basale has four ventral setae (2+2), a single lateral seta, and a short dorsal seta. The second segment has a single medial ventral seta, and the second segment single medial setae dorsally and ventrally. The length ratios of the terminal setae on the third segment are 65:100:44; the longest seta is 11%CL and 85% the length of the limb.
Comparisons. Rotundoecia is most similar to Metaconchoecia . The characters that are most clearly discriminate these two genera are summarized in Table 9 and Appendices 1–3, and illustrated in Figure 2 View FIGURE 2 .
Remarks. Iles (1953), when first describing Conchoecia teretivalvata , implied that it is conspecific with Müller’s (1894) Conchoecia rotundata short form from the Mediterranean and Müller’s coloured illustration (1894 plate 8 fig. 23) appears to support Iles’s conclusion. Gooday (1981) retained the name Conchoecia rotundata for another suitably sized and shaped Metaconchoecia species that occurs in the North Atlantic outside the Strait of Gibraltar. However, in his original description of Conchoecia rotundata from the Pacific Müller (1890) gives the length of the species to be 1.15 mm, so confirmation of the identity of both of the Mediterranean species (maybe both forms) and the Atlantic species is still needed. Sizes of specimens collected elsewhere in the tropical and temperate North Atlantic have all fallen within the size range given by Iles (1953) for R. teretivalvata . One of us (MVA) has re-examined material identified by Iles (1953) as Conchoecia teretivalvata and found in amongst typically sized specimens a few specimens that are>1.0 mm in length. Similar larger sized specimens also occurred in material collected by RRS William Scoresby close to South Georgia (Station WS 143, at 53° 58’S, 40° 30’W); these large forms may well prove to be different Rotundoecia species. Deevey (1974: Fig. 4a, c–e, j, l View FIGURE 4 ) illustrated a “larger form” of Conchoecia teretivalvata from the South Atlantic, and showed both sexes as having carapaces that are less globose than the typical form ( CH 50%CL compared with the typical CH of 55–60%CL). Thus we expect that the number of species attributed to this genus will increase as more material, particularly from high southern latitudes is examined.
Believing Iles (1953) had failed to designate either type specimens or a type locality for Conchoecia teretivalvata in his original description, so we selected a male and a female from Iles’s original Benguela Current samples to serve as comparative material for the species. Both specimens are from RSS William Scoresby station WS 977, (6 March 1950) 22°39’S, 12°16’E, and were collected from a depth of 250– 100m. These have been deposited together in the British Museum (Natural History)(♂ 2004.2552, ♀ 2004.2553). However, when going through an accumulation of material rescued from Iles’s garage two years after his death, slides of dissected male and female specimens were found which had been clearly marked as types. These slides have been deposited in the British Museum (Natural History)— holotype a male 2009.262, paratypes two females 2009.263 and 264. Entirely fortuitously the type material came from exactly the same sample from which the comparative material had been selected earlier GoogleMaps .
Distribution. Atlantic Ocean and Mediterranean Sea: occurs in the latitude range from 60ºN to 49ºS ( Müller 1894; Iles 1953; Angel 1968a, 1977, 1979, 1981b, 1983b, 1984; Angel & Fasham 1975; Deevey 1974; 1978b, Gooday 1981; Ellis 1985). Indian Ocean: known only from 4ºN, 78ºE ( Gooday 1981). Pacific Ocean: found from 22ºS to 27°S ( Poulsen 1973; Martens 1979). Deevey (1978a, 1982a, 1983) extended its range in the Pacific sector of the Southern Ocean to 65°S. In the literature the bathymetric range is given as 0–2000m, but stratified sampling in the Northeast Atlantic shows that its bathymetric range is 25– 500m.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |