
Conchoecia rudyakovi Chavtur, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4516.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:47238257-4DC1-4CF3-A07F-862FFD5E4ECF |
|
DOI |
https://doi.org/10.5281/zenodo.5959281 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87F4-1F34-2C31-10ED-FADEF54CFEF4 |
|
treatment provided by |
Plazi |
|
scientific name |
Conchoecia rudyakovi Chavtur |
| status |
sp. nov. |
Conchoecia rudyakovi Chavtur View in CoL , sp. nov.
( Figs. 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
1974 Conchoecia aff. magna Dana—Chavtur & Schornikov : 286;
1976 Conchoecia magna Dana— Chavtur: 104–105;
1977a Conchoecia magna Dana— Chavtur: 143;
1977b Conchoecia magna Dana— Chavtur: table 2;
2015 Conchoecia sp. A—Kaplun et al.: 160–161;
2015 Conchoecia sp. A—Chavtur & Bashmanov: table 2.
Etymology. The species is named in honor of the eminent ostracodologist J. A. Rudjakov.
Holotype. MIMB 1 View Materials 8344/2—adult male, length 2.42 mm, appendages mounted on a slide (18344/2–1) and valves in alcohol (18344/2–2).
Type Locality. North Pacific Ocean, Gulf of Alaska, RV Vityaz, 45 th Cruise, station 6096, sample 22, 57°20´5– 57°23´5 N and 148°35´5–148°37´2W, layer 200– 100 m, sounding 4000 m, May 8 1969.
Paratypes. RV Vityaz, 45 th Cruise, 1969 : MIMB 18344 View Materials /1—adult male, length 2.30 mm and 18344/11, 18344/ 12—adult females (2.45 and 2.40 mm), station as for holotype, sample 26, layer 300– 200 m ; MIMB 18344 View Materials /3, 18344/4, 18344/5, 18344/6, 18344/7, 18344/8 and 18344/9—adult males (lengths respectively 2.37, 2.35, 2.40, 2.38, 2.40, 2.35 and 2.30 mm) and 18344/10, 18344/13 and 18344/20—adult female (2.42, 2.40 and 2.45 mm), station and sample as for holotype . MFFT Professor Kaganovsky, 2010: MIMB 18344 View Materials /14, 18344/15, 18344/ 1618344/17—adult males (lengths respectively 2.37, 2.38, 2.30 and 2.38 mm) and 18344/18 and 18344/19, (2.50, 2.35 mm), station 65, sample 127, 43°00 N–169°00 E, layer 200–0 m, April 18.
Addition material: see Appendix.
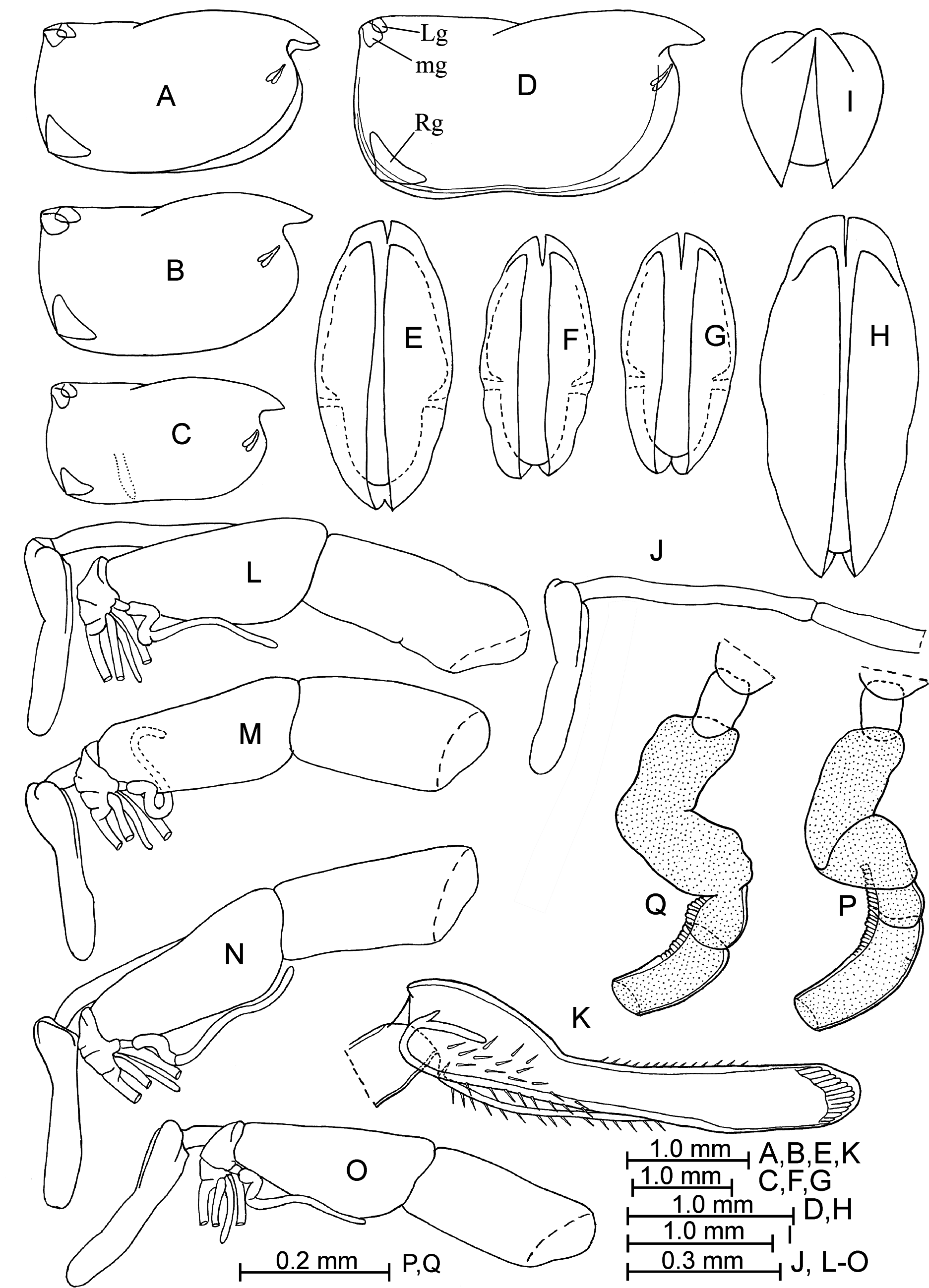
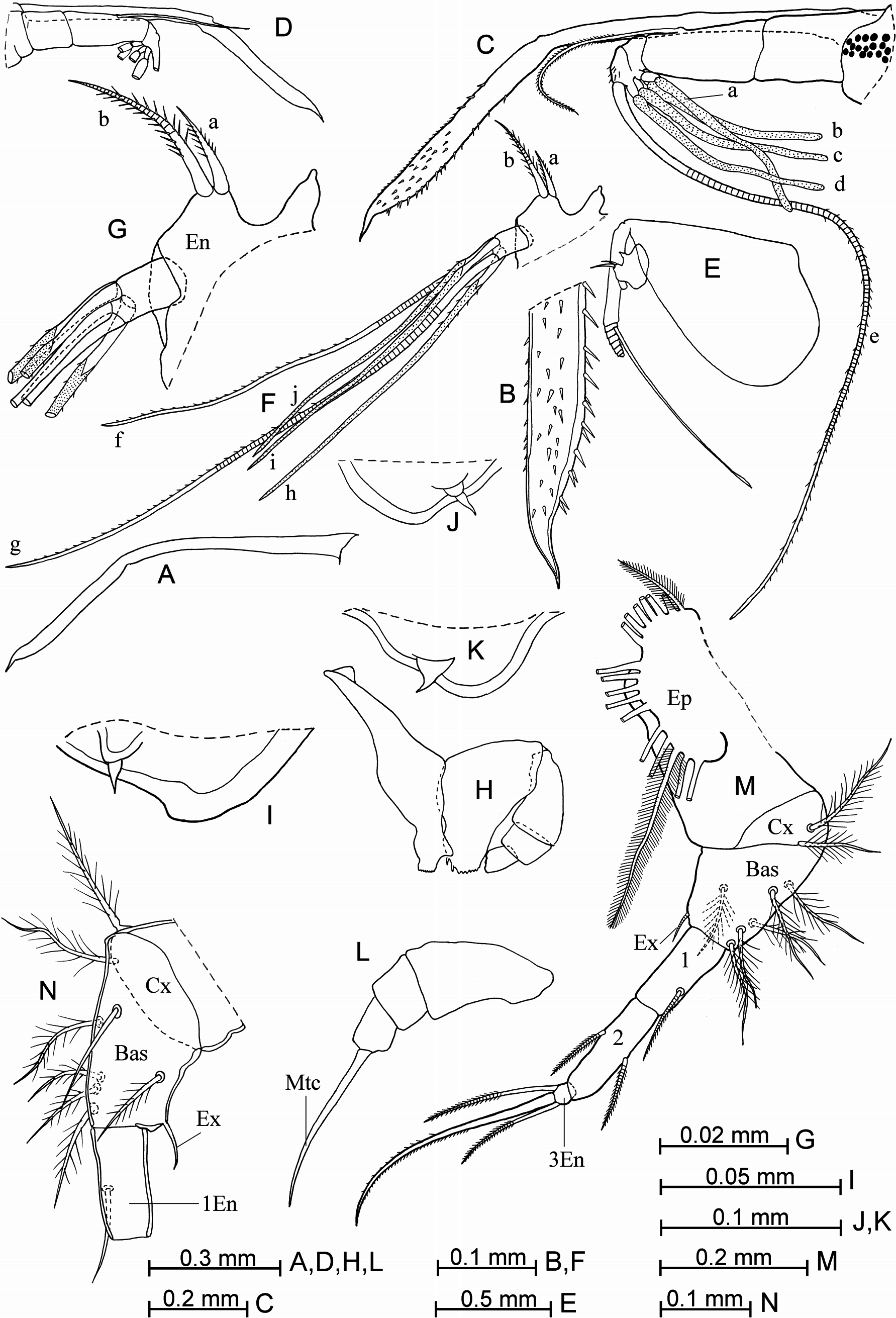
Description of adult male. Carapace ( Fig. 1 View FIGURE 1 A–I). The length is between 2.20–2.55 mm in the north-western Pacific and 2.20–2.46 mm in the north-eastern Pacific. It is rectangular. The height is 52–57% of the length and either uniform or is slightly tapered anteriorly. The asymmetrical glands and gland cells are located at the positions which are characteristic for the genus. The left asymmetrical gland is shifted anteriorly along the dorsal margin by about 4–7% of the carapace length. Sculpture consists of concentric lines parallel to margins, which is sometimes obscure.
Frontal organ ( Figs. 1 View FIGURE 1 J–O; 2A). The stem is two-segmented. The capitulum is elongated, relatively narrow, ± than length of the second segment, centrally concave dorsally, with a rounded tip, and is usually covered with medium-length spines on the lateral and ventral surface proximally and small spines on the dorsal surface distally.
First antenna ( Figs. 1 View FIGURE 1 L–Q; 2A–D). The first segment is similar in length or slightly shorter than the second segment. Seta-a just reaches or slightly extends beyond the boundary between the first and second segments. Setac is as long as or slightly longer than the combined lengths of the third, four and fifth segments. Armature of seta-e consists of a comb with about 10–12 pairs of long straight and slender spines distally and with about 12–16 alternated ones becoming a single row proximally (in total about 34–37 spines). The spines are directed basally and longer than the diameter of the seta-e. Setae-b and -d bear a few small spines and are slightly shorter than seta-e.
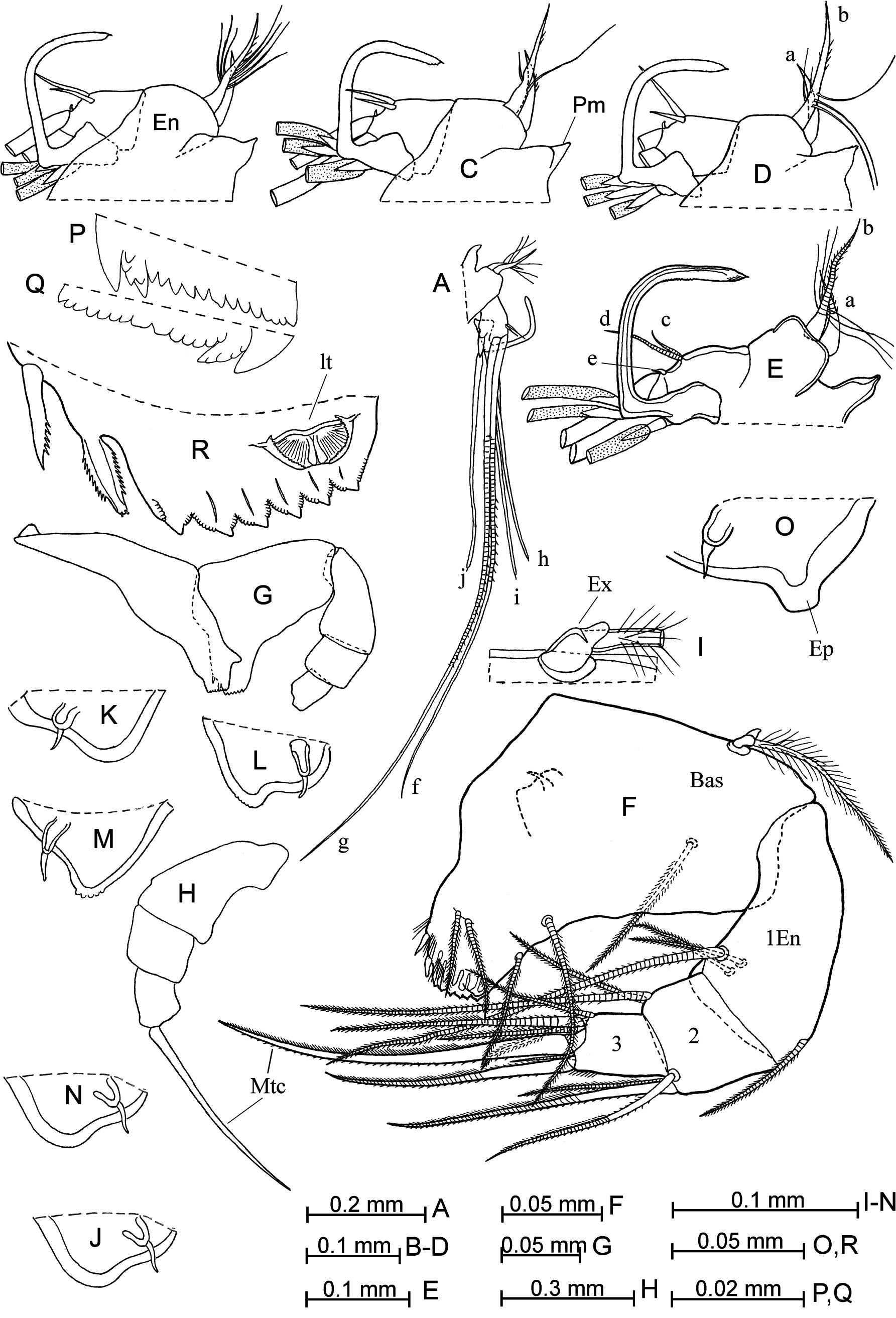
Second antenna ( Figs. 2 View FIGURE 2 E–N; 3A–E). Seta-b on the endopodite is armed with one to four long posterior and one to three medium-length anterior fine filaments. The right clasping organ is large, thin, squared and with a tip that is unswollen, pointed or spine-like. The left clasping organ is right-angled and tapers to its pointed tip. Setae-f and -g have sutures. The processus mamillaris ( Fig. 3C View FIGURE 3 ) is triangular, without or with small verruca.
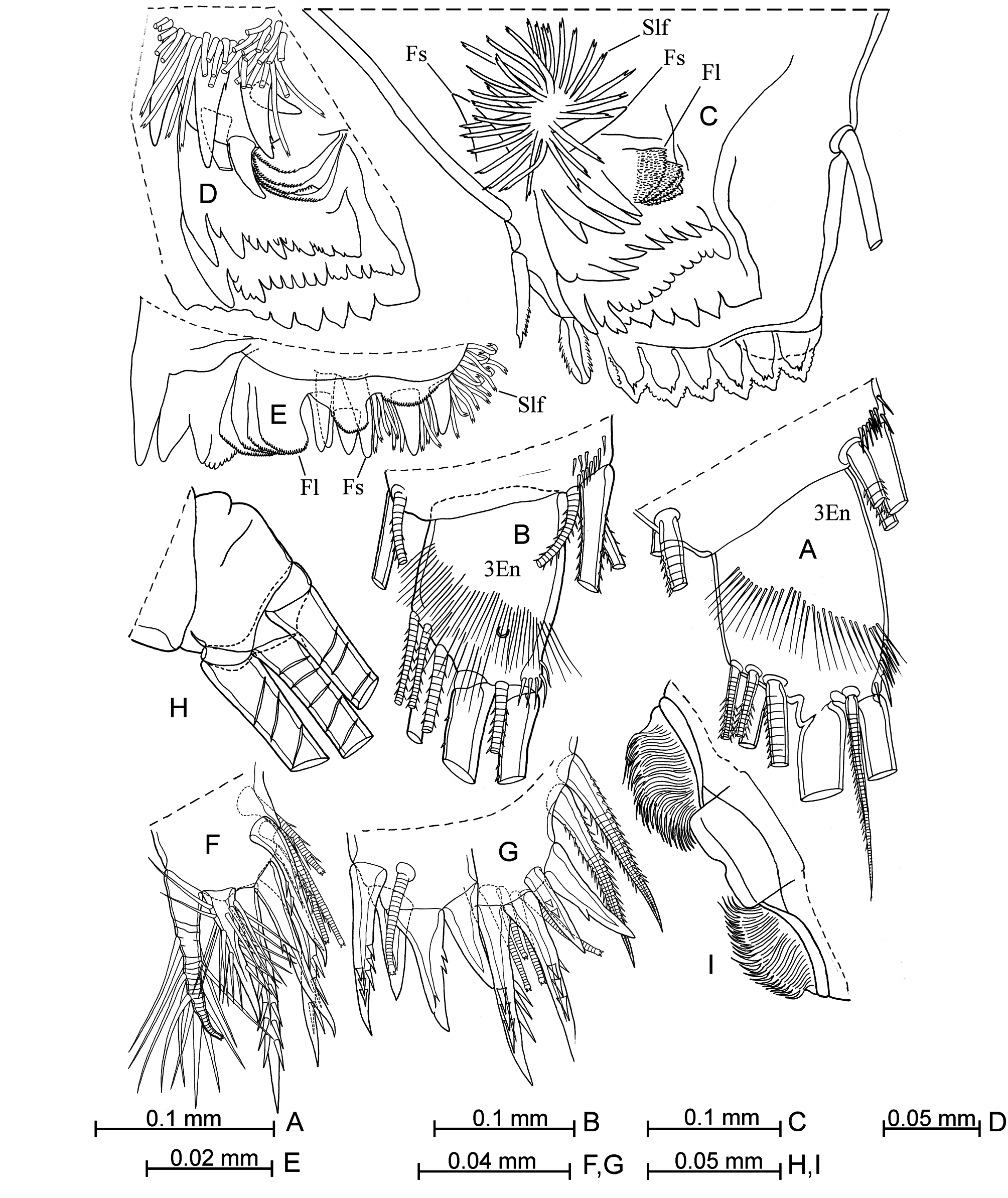
Mandible ( Figs. 3 View FIGURE 3 F–R; 4A–E; 7I). The epipodite is medium-length ( Fig. 3O View FIGURE 3 ), with noticeable verruca and a medium-length seta. The ventral margin of the first endopodite segment ( Fig. 3F View FIGURE 3 ) bears one long seta and two short setae. The disto-dorsal seta on this segment is armed with short spines. The second and third segments are covered with some stiff short spines. The main terminal claw is longer than the basale, but somewhat shorter than the endopodite, and with relatively long thin ventral spines at the middle part. The tooth edge of the coxale is armed with about ten teeth; the distal tooth-list bears 13–16 teeth and the proximal tooth-list has 8–11 teeth. The masticatory pad ( Fig. 4 View FIGURE 4 D–E) has four small rounded flaps, six flat spines and about 25–30 seta-like filaments.
Maxilla (figs. 4F, G; 5A, B). The first endopodite segment bears six anterior and three posterior setae. Along the distal edge of this segment is a row of five long and thin spines.
Fifth limb ( Fig. 5C, D View FIGURE 5 ). The basal segment has a proximal group of three or four setae ventrally, a medio-lateral group of two and a distal group of three setae. Distally, the basal segment has a long dorsal seta (vestige of the exopodite), which extends to or slightly beyond the end of the limb, and a lateral plumose seta. The first endopodite segment bears two ventral setae and one dorsal seta.
Sixth limb ( Figs. 4H View FIGURE 4 ; 6 View FIGURE 6 A–C). The coxale has two long plumose seta. The ventral margin of the basale bears five (or rarely four) setae: two long proximal setae (one of which is plumose) and three shorter distal ones. The dorsal seta (vestige of the exopodite) barely reaches or does not reach the proximal margin of the first endopodite segment. Terminal setae are subequal and plumose terminally.
Caudal furca ( Fig. 5F View FIGURE 5 ). The second claw is subequal to the height of the limb. Claws are relatively thick and all are armed with secondary spines. An unpaired seta, which is slightly longer than the eighth pair of setae, is present.
Copulatory appendage ( Fig. 6 View FIGURE 6 E–G). The limb is spindle-shaped, broadest at the middle, tapers to the end and has a rounded tip. The distal seta is very thin and with a blunt tip. The appendage is small, sclerotized and squareshaped. The limb has five to eight oblique muscle bands.
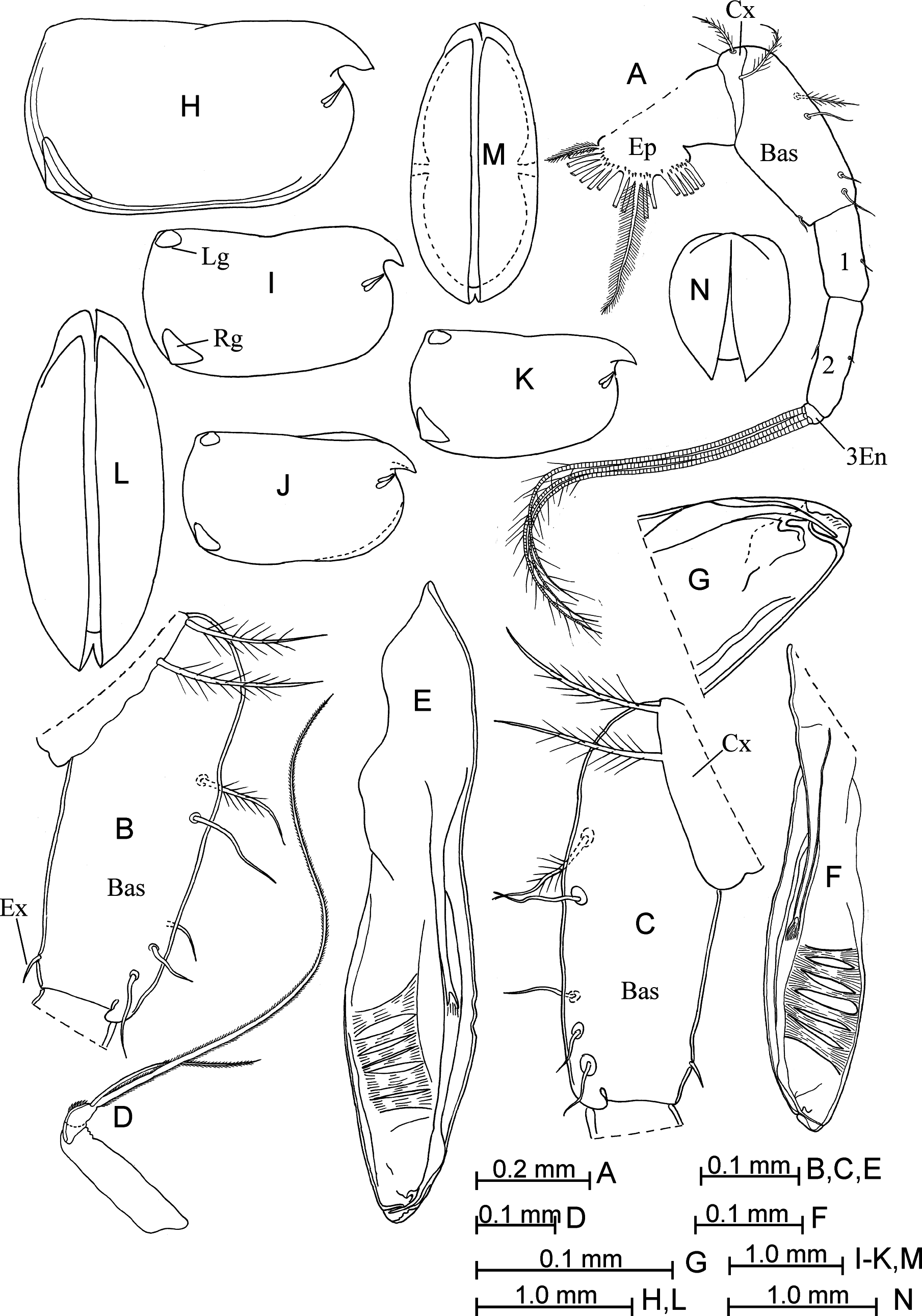
Description of adult female. Carapace ( Fig. 6 View FIGURE 6 H–N). The length is between 2.20–2.70 mm in the northwestern Pacific and 2.25–2.60 mm in the north-eastern Pacific. It is rectangular, more rounded than in the male and slightly tapered anteriorly. The height is about 55–60% of the length. The ventral margin is almost straight, and the posterior margin is arched. The locations of the glands (except of postero-medial glands) and gland cells are as in the male. The left asymmetrical gland is somewhat positioned forward along the dorsal margin by about 4–6%. Sculpture is similar to that of the male.
Frontal organ ( Fig. 7 View FIGURE 7 A–D). It is bent downward. The capitulum is long and slim, straight, fused with the stem, with a noticeably stretched and pointed downwards tip. The surface is covered with different sized spines.
C. rudyakovi C.angustihilata C. sculpta C. magna C. aff. magna C. subarcuata
Male
Carapace: Ventral margin straight straight straight straight straight concave ......continued on the next page
Length, mm 2.20–2.55 1.55–2.01 1.72–2.05 1.4–2.10 1.74–1.94 1.5–2.40 Sculpture barely visible barely visible distinct visible barely visible? barely visible
Mandible and maxilla same as in male
First antenna ( Fig. 7C, D View FIGURE 7 ). The first segment is shorter than the second segment. The dorsal seta is developed and reaches to the middle of the capitulum. Setae-a–d are relatively slim and about1/3 the length of seta-e. Seta-e is slightly tapered to a pointed tip and lacks “hairs” on the anterior margin.
Second antenna ( Fig. 7 View FIGURE 7 E–G). The first endopodite segment is about 26–27%, 35% and 45–46% the lengths of seta-g, -f and –h, respectively.
Mandible ( Fig.7 View FIGURE 7 H–L), maxilla, fifth limb and caudal furca are similar to that of the male.
Sixth limb ( Fig. 7M, N View FIGURE 7 ). The coxale has two unequal long plumose setae. The basale bears five ventral setae and one lateral plumose seta. The exopodite seta reaches 1/4–1/2 the length of the first endopodite segment.
Comparison ( Table 3). C. rudyakovi sp. nov. is closely related to C. magna and C. aff. magna , but differs by the following characteristics (in brackets for C. magna and C. aff. magna , respectively):
1 Larger sizes of the carapace, 2.20–2.55 mm in the male (1.40–2.10 and 1.74–1.94 mm) and 2.20–2.70 mm in the female (1.50–2.10 and 1.83–1.99 mm);
2 First antenna of the male: seta-a barely extends beyond the proximal end of the second segment (clearly extends beyond in both), seta-b lacks a pad (without and with pad).
3 Lengths of the paired spines on “e”-seta of the first antenna in the male are clearly greater than diameter of the seta (similar to the diameter of e-seta).
4 Processus mamillaris on the endopodite of the second antenna is triangular (tapered to rounded tip and with knob-like tip).
5 Masticatory pad on the mandible has six flat spines (four) and about 25–30 seta-like filaments (13–14 in C. magna , for C. aff. magna unknown).
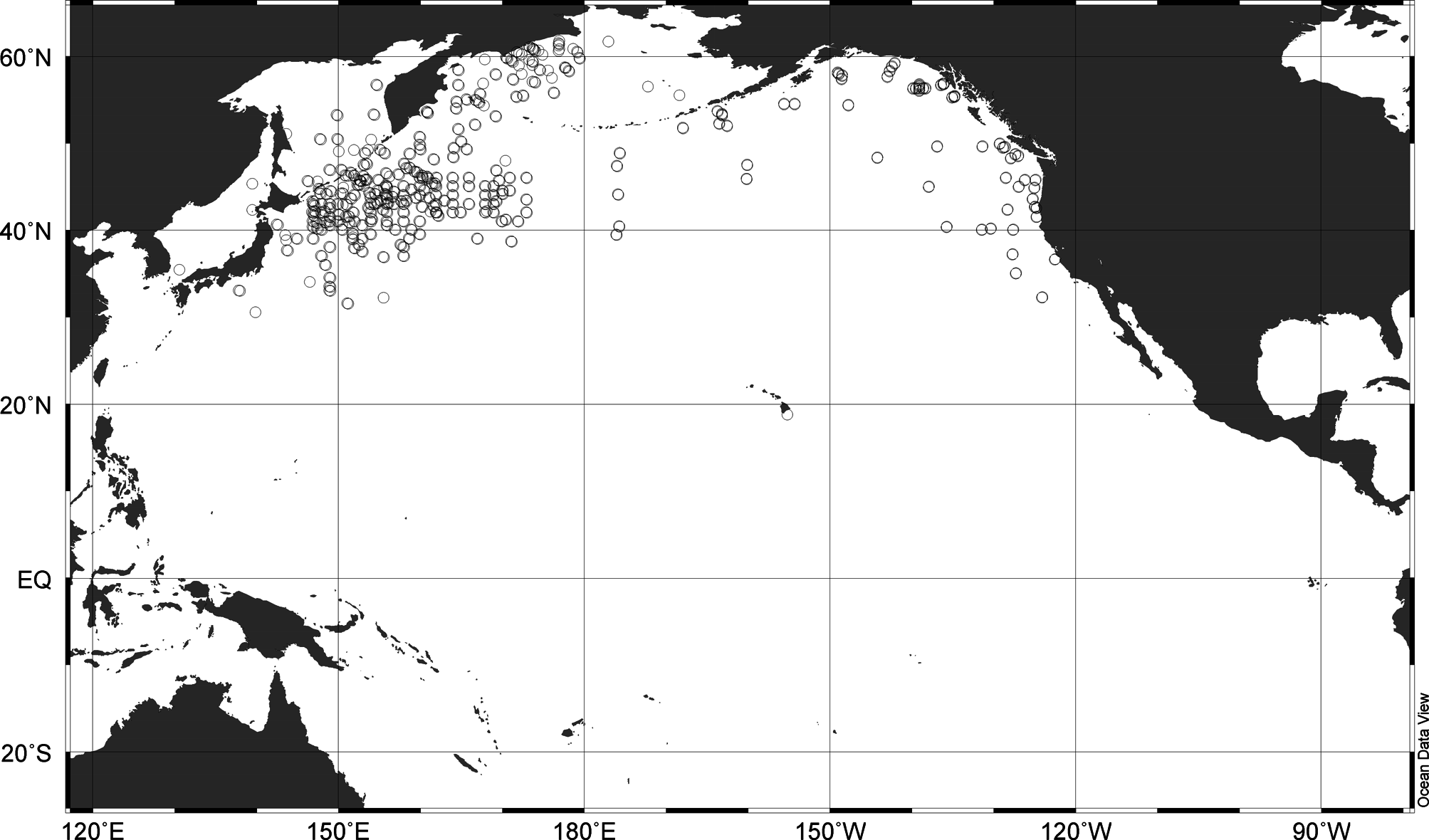
Distribution. C. rudyakovi Chavtur , sp. nov. is endemic to the waters of the sub-Arctic area and adjacent subtropical regions of the northern Pacific Ocean, ranging from 33° to 62°N (in Asian waters) and from 32°–56°N (in American waters) ( Fig. 8 View FIGURE 8 ). Thus its geographical distribution is boreal, which is in marked contrast to its congers, all of which are restricted to the tropical and subtropical waters of the World Ocean; in the North Pacific Ocean they are only encountered in the warm water zone. C. rudyakovi Chavtur , sp. nov. presumably evolved in the subtropics and then spread northwards to occupy almost the entire boreal region of the North Pacific (except for the Sea of Japan; here it has been recorded only in the waters that entered through the Sangarsky Strait from the Pacific). Thus, this species is a widely-distributed, subtropical-boreal, interzonal, epi-mesopelagic species restricted to the North Pacific. The population density, biomass and role of C. rudyakovi Chavtur , sp. nov. in plankton of the Pacific boreal waters are more significant in the American sector than that in the Asian sector ( Kaplun et al., 2015). Within its geographical range, C. rudyakovi Chavtur , sp. nov. is most abundant at 0–500 m and rarely occurs deeper than 750–1000 m (exceptionally two females were sampled from 2000–2500 m—RV Akademik Mstislav Keldysh, 1990, St. 2326, see Appendix).


| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Halocypridina |
|
Family |
|
|
SubFamily |
Conchoeciinae |
|
Tribe |
Conchoeciini |
|
Genus |