
Cheirodontinae
|
publication ID |
https://doi.org/10.5281/zenodo.10881992 |
|
DOI |
https://doi.org/10.5281/zenodo.10881801 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87B2-2A36-FFEC-FA72-F5B84DEEF7B9 |
|
treatment provided by |
Juliana |
|
scientific name |
Cheirodontinae |
| status |
|
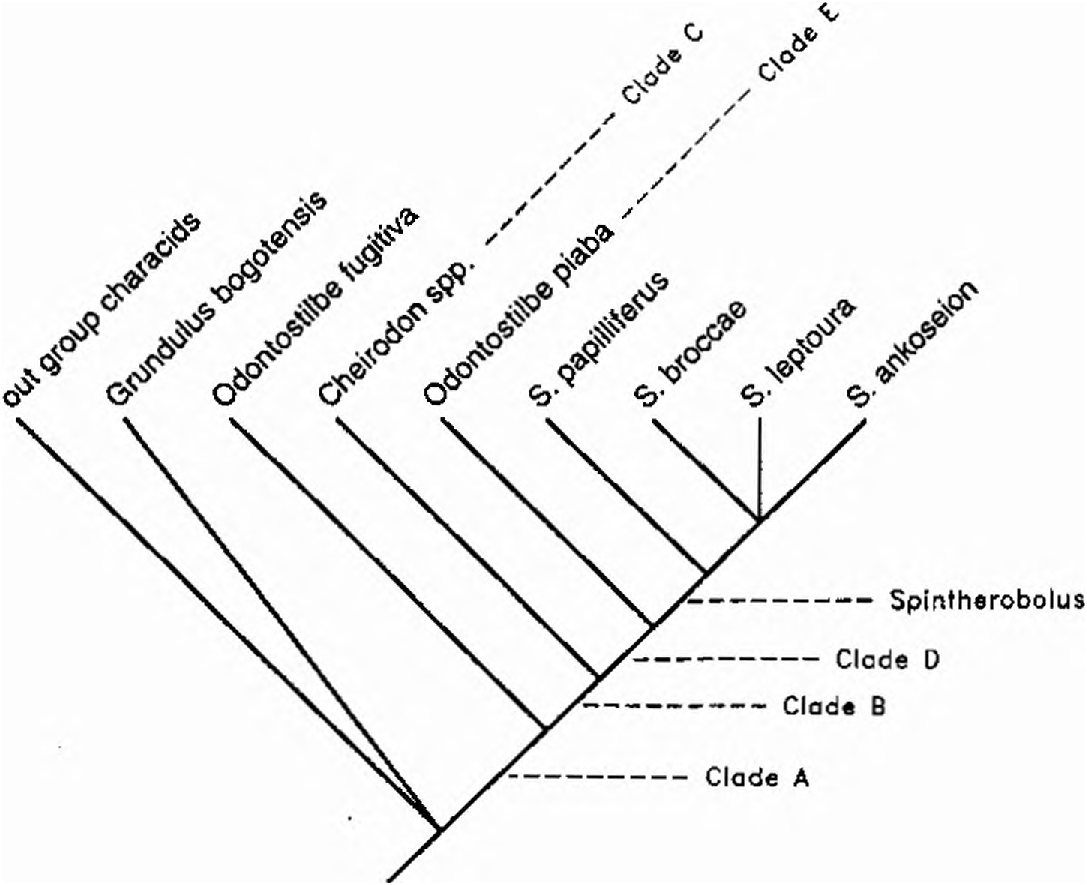
Subfamily Cheirodontinae , Clade A
Spintherobolus species belong to a restricted Cheirodontinae as diagnosed phylogenetically by Malabarba (1994; in press). For purposes of hypothesizing the phylogenetic relationships of Spintherobolus we summarize his diagnosis using the first four characters listed below. The Cheirodontinae as diagnosed by these features is equivalent to Clade A in our cladogram.
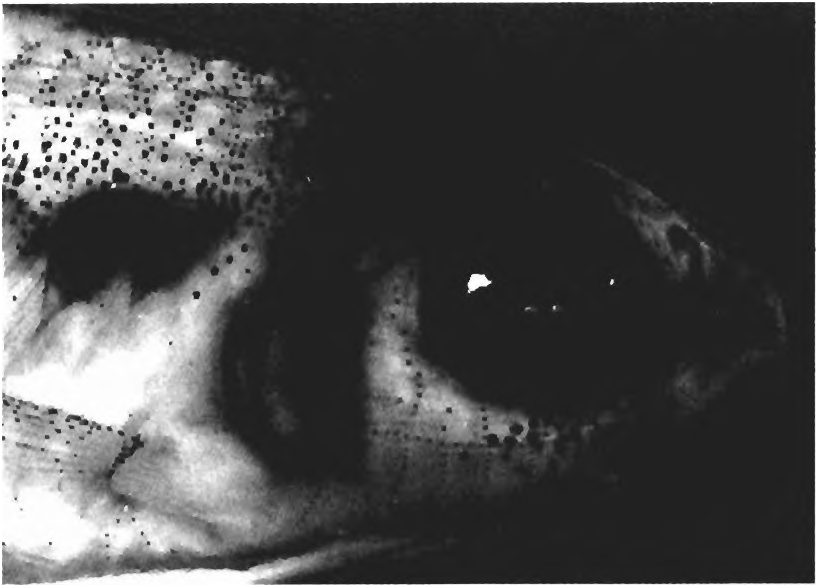
(1) A pigmented humeral spot is absent. Preserved Cheirodontines often appear to have a humeral spot but the darkened area in the humeral region is due to a muscle hiatus just lateral to the anterior portion of the swimbladder ( Fig. 4 View Fig ). The swimbladder may have dark pigment on it, but this is not the same pigment that forms the humeral spot of the epidermis of other characiforms.
Absence of a humeral spot occurs in relatively few other Characiform species, but none of these have all four of the characters listed here. This statement suggests that the lack of humeral spots in some other characiforms is homoplastic with regard to their absence in the Cheirodontinae . At this time we are unable to provide an analysis of the cladistic significance of the humeral spot and its absence in other characiforms because the phylogenetic significance of the distribution of its presence or absence is unstudied and unknown in nearly all of them. We can only suggest that the loss of this character occurred independently in certain outgroup characids.
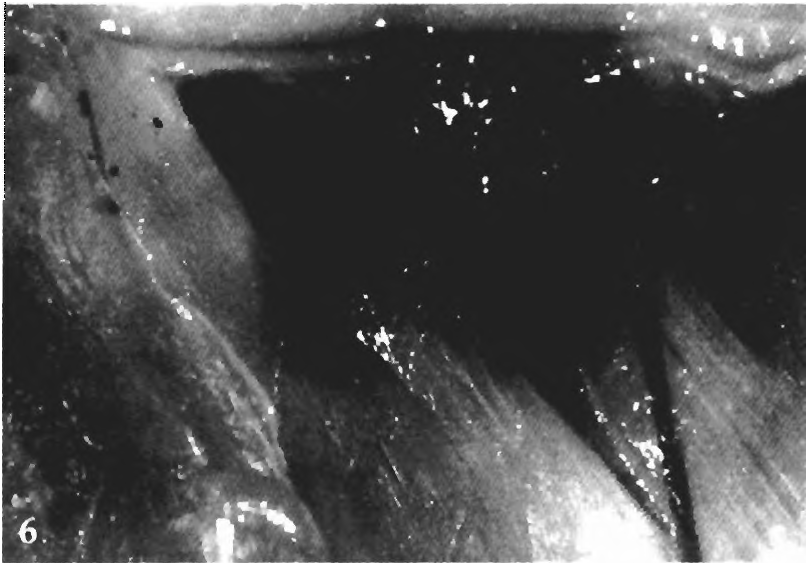
(2) A characteristic hiatus occurs in musculature of the body wall in the area of the swimbladder between the first and second pleural ribs, exposing the swimbladder ( Figs. 4-6 View Fig View Fig View Fig ). This hiatus is not a simple derived absence of musculature tissue, but a derived and organized feature of the body wall presumably making it more efficient in transmitting sound from the environment to the swimbladder, perhaps by avoiding interference from active muscle tissue. This hiatus is an organ called the pseudotympanum because the membranous wall of the swimbladder is exposed to fluid and some fat in the muscular hiatus. The absence of interference with sound transmission from active muscle tissue may increase the efficiency of that transmission from water external to the skin and scales to the wall of the swimbladder. From there sound is transmitted to the inner ear through the intermediate contact between the swimbladder wall and the Weberian apparatus. Also the absence of muscle tissue against the membranous swimbladder wall may allow that wall more freedom to respond to sound waves. Note: The species of Spintherobolus have two rather than one pseudotympanum and their posterior pseudotympanum (between the first and second pleural ribs) is homologous to that in all other species of the Cheirodontinae ; see more detailed description in synapomorphy 21 below for Spintherobolus .
Among other characiforms, a pseudotympanum of different structure and presumably not homologous with that found in the Cheirodontinae is also found in the species of the Characinae. The implications of this are discussed by Malabarba (in press).
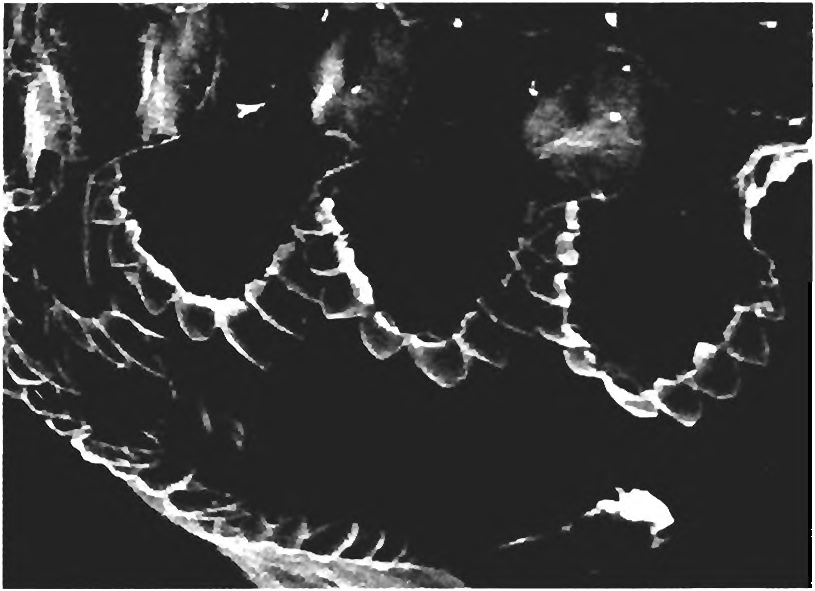
(3) Cheirodontine teeth have a proximal peduncle or pedicle and an expanded and compressed distal region bordered by a few to many cusps ( Fig. 7 View Fig ). Note: In the case of the species of Spintherobolus the modification of this flattened, expanded distal portion into an essentially conical tooth is a secondary alteration according to the hypothesis of relationships presented here; see also apomorphy 3R below. Note that not all of these teeth in Spintherobolus are conical.
Somewhat similar teeth occur in Piabucus and Iguanodectes , genera referred to the tribe Iguanodectini of the Tetragonopterinae by Géry (1977). Even when these genera are entered in our analysis and their teeth are coded the same as those of the Cheirodontinae , these two genera are ex cluded from that subfamily in the most parsimonious cladogram.
(4) All Cheirodontines have a single series of teeth on the premaxilla. This feature is discussed by Malabarba (in press) but it is not restricted to the Cheirodontinae and has appeared convergently many times in the Characidae and some other characiforms.
The Cheirodontinae consists of several major clades ( Malabarba, 1994; in press). We treat only one of these, Clade B containing Spintherobolus , in detail in our cladogram ( Fig. 3 View Fig ). We defer treatment of the other cheirodontine clades to Malabarba (in press). The species of Spintherobolus form a subgroup of clade B and share several derived features of the ventral procurrent caudal-fin rays not found in other characiforms, including other clade A Cheirodontines. Other Clade A Cheirodontines are represented in Figure 3 View Fig by Odontostilbe fugitiva Cope , the type species of Odontostilbe .
In addition to the species of Spintherobolus Clade B Cheirodontines include the species of Cheirodon (sensu Casciotta et al., 1992) and are here labeled as Clade C in Figure 3 View Fig . Some other species at present placed in the polyphyletic and paraphyletic Odontostilbe are here placed in Clade E in our cladogram and the earliest described species of this clade, Odontostilbe piaba, represents the included species our Figure 3 View Fig .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.