
Spintherobolus papilliferus Eigenmann, 1911
|
publication ID |
https://doi.org/10.5281/zenodo.10881992 |
|
DOI |
https://doi.org/10.5281/zenodo.10881809 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87B2-2A20-FFFC-FF53-F9594FA8F9E9 |
|
treatment provided by |
Juliana |
|
scientific name |
Spintherobolus papilliferus Eigenmann |
| status |
|
Spintherobolus papilliferus Eigenmann View in CoL
( Figs. 1 View Fig , 26-28 View Fig View Fig View Fig )
Spintherobolus papilliferus Eigenmann, 1911: 167 View in CoL , pl. 5 figs. 1 -4 (new species description, type locality: Alto da Serra , São Paulo, Brazil, holotype: CM 2582 , now FMNH 104802 ; 4 paratypes: CM 2583 , now FMNH 54918 and CAS[SU] 17520 )
Specimens examined. All from Brazil, São Paulo, upper rio Tietê drainage. FMNH 104802 , holotype, immature; 34.1 mm SL; FMNH 54918 , 3 paratypes, immature, 20.2,30.5, 30.7 mm SL; CAS-SU 17520 , 1 paratype, juv., 28.5 mm SL (note: FMNH specimens mixed and holotype not separately labeled; see Discussion section below); Alto da Serra, 4 Aug 1908, J. D. Haseman. - MZUSP 51022,17 , all immature, 16.7-24.6 mm SL; Paranapiacaba and Campo Grande; 24 Mar 1967, S. P. Werner. - MZUSP 49408 , 1 male 57.8 mm SL, 2 females, 50.3-50.4 mm SL, 1 female, c&s, 45.0 mm SL; “last creek on road to Paranapiacaba ”; 20 Mar 1980, R. M. C. Castro. - MZUSP 51021 , 1, female, 60.8 mm SL; rio Ipiranga ; January 1929, H. Luederwöldt. - MNRJ 6260 , 1, female, 35.8 mm SL (same specimen in Travassos, 1953: 506, fig. 1). - MNRJ 4237 , 1 male, 34.5 mm SL, 4 females, 32.8- 36.5 mm SL; rio Ipiranga; date unknown but probably before 1931, A. Couto de Magalhães.
Diagnosis. The following five autapomorphies (20, state 3; 27, state 2; 29; 30; 31) distinguish S. papilliferus from all other species of the genus. Also three anal-fin reversals (10 R, 11 R, 12 R) help distinguish this species.
(20, state 3) Branched anal-fin rays 9-10. This is the lowest count for this fin known for the Cheirodontinae . The other three species of Spintherobolus have 11-16 branched anal-fin rays.
(27, state 2) The eyes of S. papilliferus (18.4- 26.0, x = 21.3 % HL, n = 31) are the smallest of the four species of Spintherobolus . The eyes of Spintherobolus ankoseion (24.6-30.1, x = 27.6 %, n = 22), those of S. broccae (24.5-32.7, x = 28.7 %, n = 70), and those of S. leptoura (23.2-30.3, x = 27.2 %, n = 18) are significantly larger relative to head length than those of S. papilliferus and are not significantly different from each other. See also discussion of apomorphy 27, state 1, above.
(29) The caudal peduncle length of S. papilliferus is long, being 23.8-24.6 % SL (x = 24.2 %) in males and 21.3-27.0 % SL (x = 23.9 %) in females. The other three species have caudal peduncle lengths between 11.5-21.5 % SL. This demonstrates that almost no overlap occurs between S. papilliferus and the other species of Spintherobolus except in an occasional female. In species of outgroup clade E the caudal peduncle length varies between 11.0-19.6 % SL and the ranges of the means extend from 14.3-15.2 % SL.
(30) Large adultsize. Mature adults of S. papilliferus reach at least 60.8 mm SL (a mature female) and 57.8 mm SL (a mature male). Another male 34.5 mm SL had developing secondary sex ual characters. Specimens of S. broccae , S. leptoura and S. ankoseion are fully mature by at least 18 mm SL and adult males were recorded to reach 23.1 mm SL ( S. ankoseion ) while adult females reach 27.9 mm SL ( S. ankoseion ). Adults of outgroup clade E species are only known to range approximately between 20-40 mm SL.
(31) Spintherobolus papilliferus has a high ver tebral count of 35-36. Compare vertebral counts of the species of Spintherobolus in Figure 21 View Fig . Species of its sister subgroup b have 32-34 vertebrae and species in outgroup clade E are known to have 32-34 vertebrae.
(10 R) The anal-fin rays of adult males are not expanded in a sagittal plane. See apomorphy 10, clade D, for outgroup condition and its taxonomic distribution.
(11 R) Segments of anal-fin rays of adult males are not fused. See apomorphy 10, clade D, for outgroup state and its taxonomic distribution.
(12 R) An anterior extension of the proximal end of the lepidotrichia of the anal-fin rays of males is absent. See apomorphy 12, clade D, for the outgroup state.
Description. Morphometric data are summarized in Table 2 View Table 2 . The reduced anal fin and lack of an adipose fin in part give this species a rather unusual body shape for a characid. Body compressed, moderately elongate. Predorsal body profile convex with short concavity at nape followed by relatively steep convexity. This more strongly developed in large specimens (above 50 mm SL). Body profile gently convex behind nape to dorsal-fin origin and along dorsal-fin base. Body profile nearly straight between terminal base of dorsal-fin and caudal-fin origin. Ventral profile of head gently convex. Belly profile between pectoral and pelvic fins straight or slightly convex. Profile between pelvic-fin origin and anal-fin origin slightly concave. Body profile along anal-fin base nearly straight or slightly concave. Caudal peduncle elongate with dorsal and ventral profiles nearly straight.
Snout prominent, blunt, and rounded. Dorsal profile ofhead straightto slightly concave. Mouth terminal, angled posteroventrally. Maxilla short, not reaching line drawn vertically through anterior margin of eye. Eyes smaller than snout length.
Dorsal-fin rays, ii, 9 in all specimens. Dorsal-fin origin near midbody distance. Adipose fin absent.
Anal-fin rays iii, 10 (anterior unbranched rays iii in 29 specimens and 2 specimens with a fourth vestigial anterior element detectable in radiographs; 29 specimens with 9 branched anal-fin rays; holotype and one other specimen with 10 branched rays; Fig. 20 View Fig ). Profile of anal fin rounded, with tip of all branched rays ending at more or less vertical imaginary line along posterior border of anal-fin base. Anal fin without hooks in two males presumably large enough to be mature. Only one fully adult male examined and its anal-fin rays longer than those of females. Anal-fin origin at vertical line drawn through origin of posteriormost dorsal-fin ray.
Pectoral-fin rays i, 13 (branched rays 10-13, x = 11.1, n = 20). Pectoral fin large, rounded, posterior tips of longest pectoral-fin rays extend to nearly pelvic-fin origin.
Pelvic-fin rays i,6 (branched rays 5-6, x = 5.8, n = 22). Pelvic fins of adult male reaching beyond anal-fin origin but not reaching that fin’s origin in females. Largest male with very short, truncated hooks located on ventromedial surface of pelvic-fin rays. These occur along one third of unbranched ray and about half of proximal length of first four branched rays.
Principal caudal-fin rays 10/ 9 (9/ 9 in one specimen, x = 18.9, n = 28). Dorsal procurrent caudal-fin rays 14 (range = 11- 15, x = 13.0, n = 24). Ventral procurrent caudal-fin rays 15 (range = 12-17, x = 14.2, n = 26; Fig. 23 View Fig ). Ventral procurrent caudal-fin rays of males strong and well-developed, with proximal extremities fused, slab shaped and expanded in sagittal plane. Last five caudal vertebrae (including posterior half centrum) supporting caudal fin and procurrent rays, with enlarged and modified hemal spines. These five caudal vertebrae thus equal precaudal vertebrae by definition. Anteriormost 10 ventral procurrent fin rays of mature males fused, with lateral processes inserted among muscle fibers and skin of this region.
Scales cycloid. Lateral line with 3-6 perforated scales (x = 4.6, n = 19). Scales in longitudinal series 34-36 (x = 35.4, n = 19; Fig. 22 View Fig ). Scale rows between dorsal-fin origin and longitudinal series (series continuous with perforated scales) 6-7 (x = 6.3, n = 19) and between this scale series and pelvic-fin origin, 5-6 (x = 5.6, n = 19). Predorsal scales from occipital process to dorsal-fin origin 12-15, irregular and difficult to count accurately, with 12-15 scales. Scale rows around caudal peduncle, 18 in all specimens. No modified scales on caudal-fin or scale sheath along anal-fin base.
All jaw teeth with expanded basal pedicle and more or less conical along their length, but with slightly expanded distal tip. Distal conical part of dentary teeth with small medial and small lateral cusps. Middle cusp relatively large, elongate and slender. Anterior premaxillary teeth also with one cusp on each side, these extremely reduced and almost imperceptible. Maxillary teeth conical with somewhat laterally expanded distal conical portion. There are 8 premaxillary, 7 max illary and 12 dentary teeth in one cleared and stained specimen, 45.0 mm SL.
Vertebrae 35-36 (x = 35.1, n = 30; Fig. 21 View Fig ).Branchiostegal rays 4. Two rays attached nearly together on midventral surface of anterior ceratohyal, one on posteroventral lateral surface of anterior ceratohyal, and one on the mid-ventrolateral surface of posterior ceratohyal, in one cleared and stained specimen.
Color in alcohol. The color description of preserved adults given below was taken from spec imens collected within the last 17 years. The color pattern of the juvenile types of S. papilliferus are somewhat faded but essentially the same as that of the more recently collected adult specimens. See Figures 1,10,11,26,27, & 28 for the preserved color patterns of adult males and females. The dorsal and lateral parts of the body are pale to medium brown, and pale yellowish brown ventrally. No obvious discrete lateral body stripes, bands, or blotches are present. In the photographs of a male and female, there appears to be a dark humeral spot just posterior to the more dorsal region of the opercular flap. This is not due to an increase number or size of the dark chromatophores. It is rather the area of the anterior and posterior pseudotympanums where the muscles of the lateral body wall are thin or absent. Black pigment coating the swimbladder can be seen through the skin and appears dark in the photograph.
The scale borders, especially their posterior borders are covered by a relatively wide band of dark chromatophores, producing a reticulate pattern over the body. These chromatophores are darker and perhaps more numerous dorsally making the belly region relatively pale and the back region relatively dark. What looks like a caudal spot in Figure 27 View Fig of the female is an artifact of lighting in taking the photograph. No area of any scale is free of dark chromatophores and the central region of all scales thus appears pale brown.
The pectoral and pelvic fins of the female are hyaline with a few scattered dark chromatophores. The pectoral fins of the male are relatively dark, with numerous dark chromatophores scattered over both the dorsal and ventral surfac es. The male’s pelvic fins are lighter, much like those of the female. The dorsal fin has a scatter ing of dark chromatophores, which are more concentrated in the mid-dorsal region of the fin. The anal and caudal fins are hyaline with a thin scattering of small dark chromatophores over the rays and membranes. The patterns appearing in the caudal- and anal-fin photographs are due to the relative thickness of the fin and how it reflects and refracts light, not to pigment patterns. The head is pale brown around the mouth and on the ventral surface of the snout, jaws, and opercular region. The dorsal surface of the snout, between the eyes, the dorsum of the cranium and the nape are all dark brown. The head area posterior to the circumorbitals and extending ventrally from the parietal region across the dorsal opercular region is pale brown in both sexes.
Color in life unknown.
Sexual dimorphism. Males and females are not easily distinguished externally, having a similar body shape and color pattern ( Figs. 1 View Fig , 26 View Fig , 27 View Fig ). External sexually dimorphic features are not ex tensively developed. The ventral procurrent caudal-fin rays of males are fused to each other, more developed than those of females, and have lateral processes. These rays are embedded deep into the scales and skin in this species and can be only identified after dissection or osteological preparation. The presence of small hooks on the pelvic fins is the clearest external sexually dimor phic feature found in this species. The pelvic fin reaches the anal-fin origin in males, but not in females.
Status of the holotype. Eigenmann (1911: 167) described S. papilliferus based on five specimens, the holotype, CM 3582, and four paratypes (designated cotypes by Eigenmann), CM 3583. We found five specimens labeled as paratypes, four in FMNH 54918, and one in CAS-SU 17520. We find that according to the labels in both jars FMNH 54918 originally contained five specimens, and one was subsequently transferred to SU 17520. However, Ibarra & Stewart (1987: 81) stated that the holotype was missing and five paratypes were present. Since they do not give FMNH museum numbers we are uncertain of the meaning of their statement. However, the holotype must be one of the five specimens present in the CAS and FMNH collections. Eigenmann gave the total length of the holotype as 41 mm and the paratypes as ranging from 25-39 mm in total length. Unfortunately all the specimens now have damaged caudal fins, but we identify the largest in the FMNH collection as the holotype ( Fig. 28 View Fig ). Its general body shape and color pattern also fit well with that published in Eigenmann (1911: pl. 5 fig.l; 1915: pl.3 fig. 1).
Conservation note. Menezes and Weitzman are extensively documenting the location of the Alto da Serra type locality near the head waters of the rio Grande da Serra, a tributary of the rio Tietê in the vicinity of Campo Grande and Paranapiacaba, State of Sao Paulo. The known collecting sites for this species have been altered by pollution and commercial use, especially that of the rio Ipiranga in the city of São Paulo where the fish is apparently extinct. We have examined one large female 60.8 mm SL ( MZUSP 51021; Figs. 11 View Fig , 26) collected in 1929 from this locality that has grooves on the top of the head that are deeper than any other specimen examined. This specimen also has 33 vertebrae rather than the 35-36 found in all other specimens of S. papilliferus. The young specimens we have from rio Ipiranga have the usual number of vertebrae found in S. papilliferus and because they are young, their cephalic grooves are not well-developed. The absence of more adult, mature specimens of S. papilliferus from the rio Ipiranga locality and with only one mature male and three mature females of this species from the rio Grande da Serra locality prevent us from investigating further the possibility that the large specimen from rio Ipiranga might be a different species.
At present S. papilliferus may survive only in the rio Grande da Serra, near Paranapiacaba and Campo Grande. This area is next to the storage and transfer yards of the Rede Ferroviária Feder al S. A (Brás) where it crosses the summit of the Serra do Mar (locally called the Serra de Paranapiacaba) between the cities of São Paulo and Santos. This area lies near the Reserva Biológica de Paranapiacaba where S. papilliferus may still ex ist. No extensive search has been made for this species in the Paranapiacaba region, but recent collection trips to the immediate vicinity ofParanapiacaba and Campo Grande failed to produce this fish.



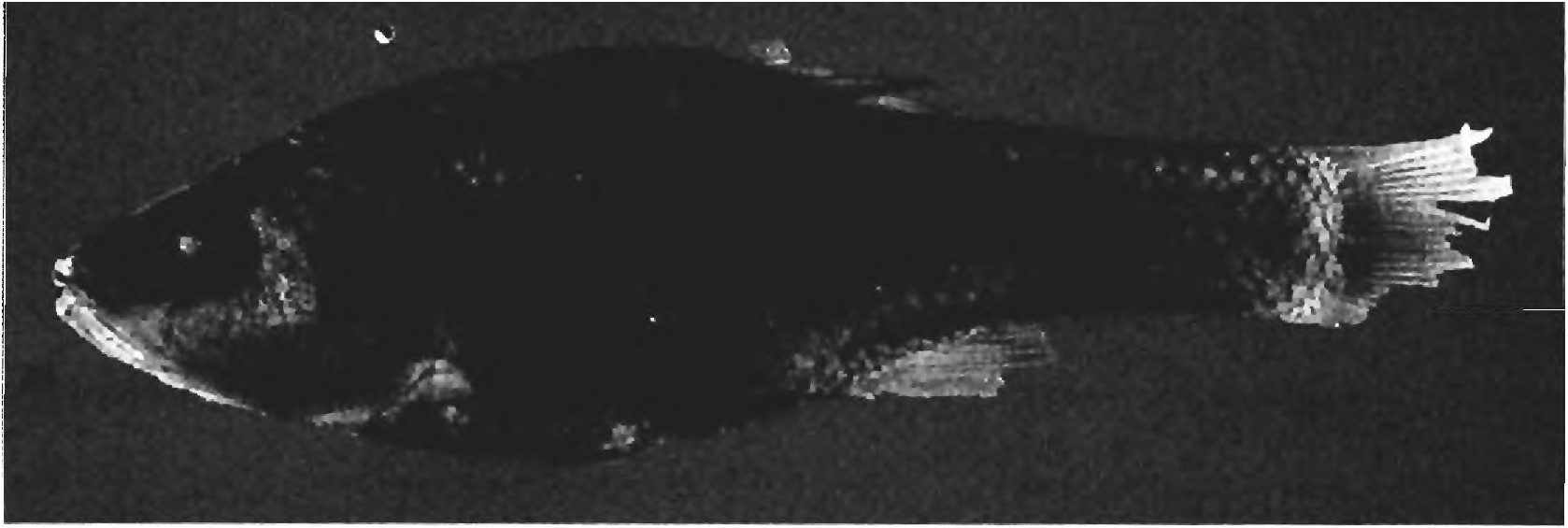
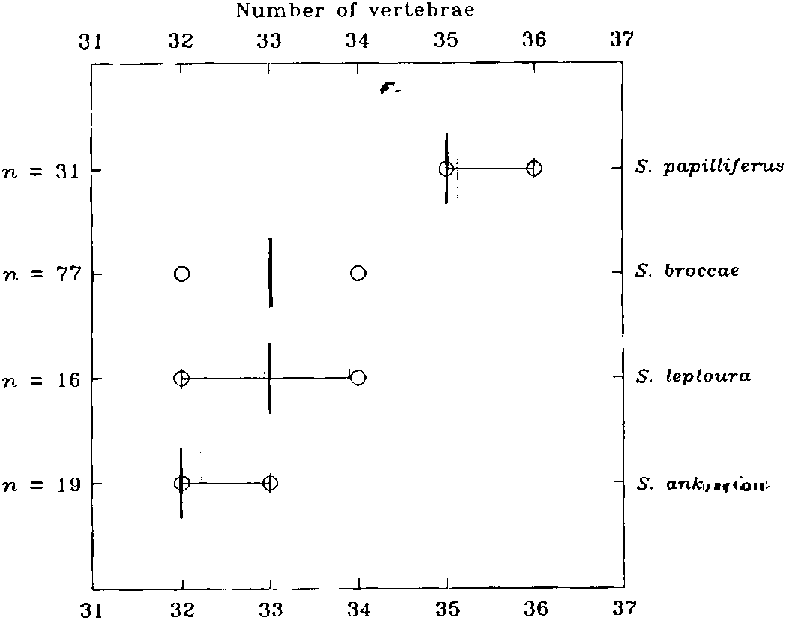
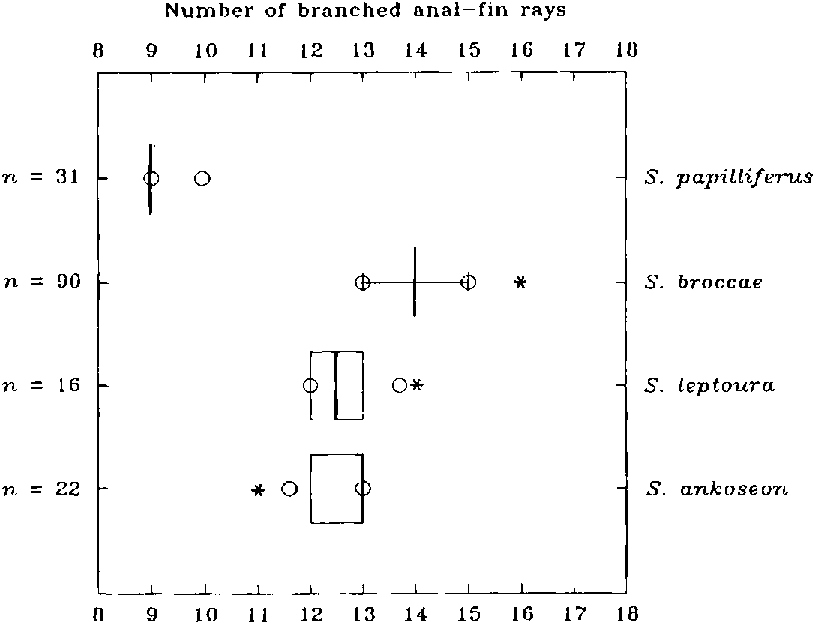
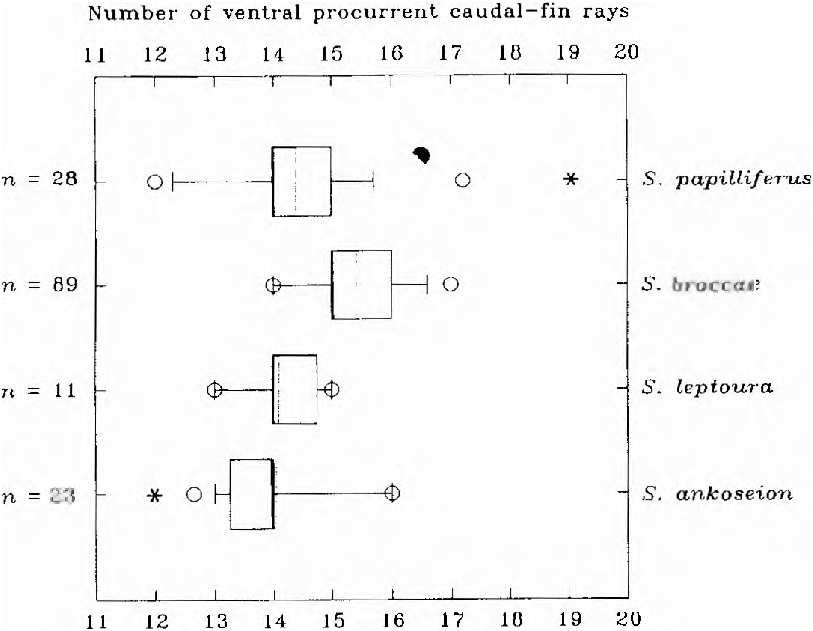
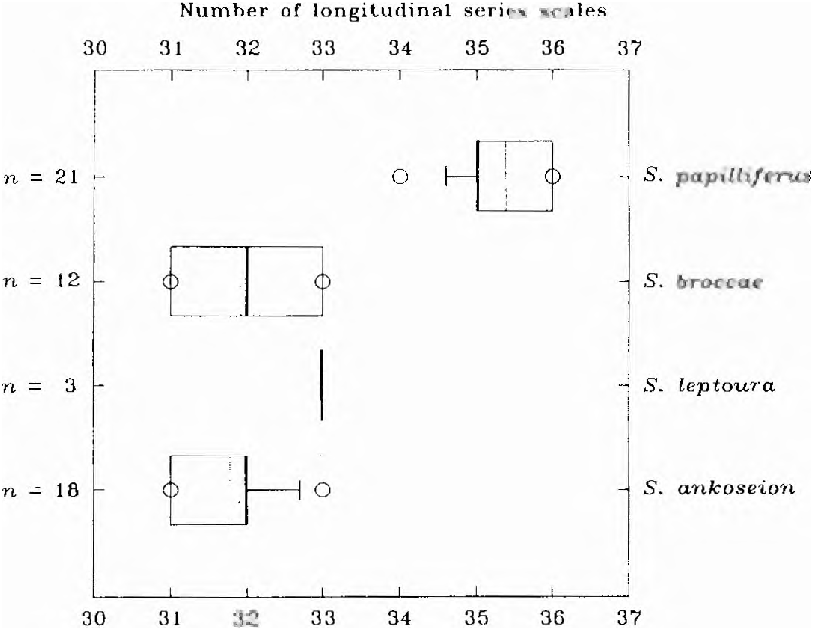

Table 2. Morphometries of Spintherobolus papilliferus. Table includes measurements of all examined specimens. Values for the holotype are given separately.
| holotype | males | females | |||||||
|---|---|---|---|---|---|---|---|---|---|
| n | low | high | mean | n | low | high | mean | ||
| Standard length [mm] | 34.1 | 2 | 34.5 | 57.8 | 46.2 | 30 | 16.7 | 60.8 | 27.7 |
| Percentage of standard length | |||||||||
| Snout to anal-fin origin | 61.0 | 2 | 61.7 | 63.3 | 62.5 | 30 | 58.7 | 67.3 | 62.2 |
| Snout to dorsal-fin origin | 54.3 | 2 | 51.9 | 52.2 | 52.0 | 29 | 50.9 | 58.2 | 54.4 |
| Snout to pelvic-fin origin | 47.5 | 2 | 42.0 | 47.4 | 44.7 | 30 | 44.1 | 50.0 | 46.9 |
| Dorsal-fin base length | 9.4 | 2 | 9.6 | 12.3 | 10.9 | 29 | 9.4 | 12.4 | 11.1 |
| Anal-fin base length | 11.4 | 2 | 12.5 | 12.6 | 12.5 | 30 | 10.4 | 14.2 | 12.5 |
| Caudal peduncle length | 27.0 | 2 | 23.8 | 24.6 | 24.2 | 30 | 21.3 | 27.0 | 23.9 |
| Caudal peduncle depth | 12.9 | 2 | 13.3 | 15.4 | 14.4 | 30 | 11.4 | 15.8 | 13.3 |
| Depth at dorsal-fin origin | 29.6 | 2 | 28.7 | 31.8 | 30.3 | 29 | 26.4 | 32.2 | 28.8 |
| Dorsal-fin length | - | 2 | 22.5 | 24.3 | 23.4 | 27 | 17.0 | 27.4 | 23.1 |
| Pelvic-fin length | 12.9 | 2 | 14.5 | 15.9 | 15.2 | 29 | 11.4 | 15.9 | 13.3 |
| Pectoral-fin length | 10.9 | 2 | 15.7 | 16.6 | 16.1 | 29 | 10.9 | 16.5 | 14.0 |
| Bony head length | 29.3 | 2 | 28.7 | 29.8 | 29.2 | 30 | 26.8 | 32.0 | 30.2 |
| Percentage of head length | |||||||||
| Snout length | 24.0 | 2 | 24.4 | 25.3 | 24.8 | 29 | 18.3 | 29.4 | 23.3 |
| Upper jaw length | 20.0 | 2 | 20.2 | 20.9 | 20.6 | 29 | 16.0 | 23.0 | 19.5 |
| Horizontal eye diameter | 26.0 | 2 | 19.2 | 20.2 | 19.7 | 29 | 18.4 | 26.0 | 21.4 |
| Least interorbital width | 25.0 | 2 | 31.3 | 31.4 | 31.4 | 29 | 22.4 | 32.0 | 26.9 |
| FMNH |
USA, Illinois, Chicago, Field Museum of Natural History (also used by Finnish Museum of Natural History) |
| CAS |
USA, California, San Francisco, California Academy of Sciences |
| MZUSP |
MZUSP |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cheirodontinae |
|
Tribe |
Cheirodontini |
|
Genus |
Spintherobolus papilliferus Eigenmann
| Weitzman, Stanley H. & Malabarba, Luiz R. 1999 |
Spintherobolus papilliferus
| Eigenmann, C. H. 1911: 167 |