
Brachypsectra cleidecostae, Lawrence & Monteith & Reid, 2020
|
publication ID |
https://doi.org/10.11606/1807-0205/2020.60.special-issue.02 |
|
publication LSID |
lsid:zoobank.org:pub:42B41A88-EC52-426A-9E66-5F15CB20CF1E |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87AB-7B65-4F73-69E5-110A37862662 |
|
treatment provided by |
Carolina |
|
scientific name |
Brachypsectra cleidecostae |
| status |
sp. nov. |
Brachypsectra cleidecostae sp. nov.
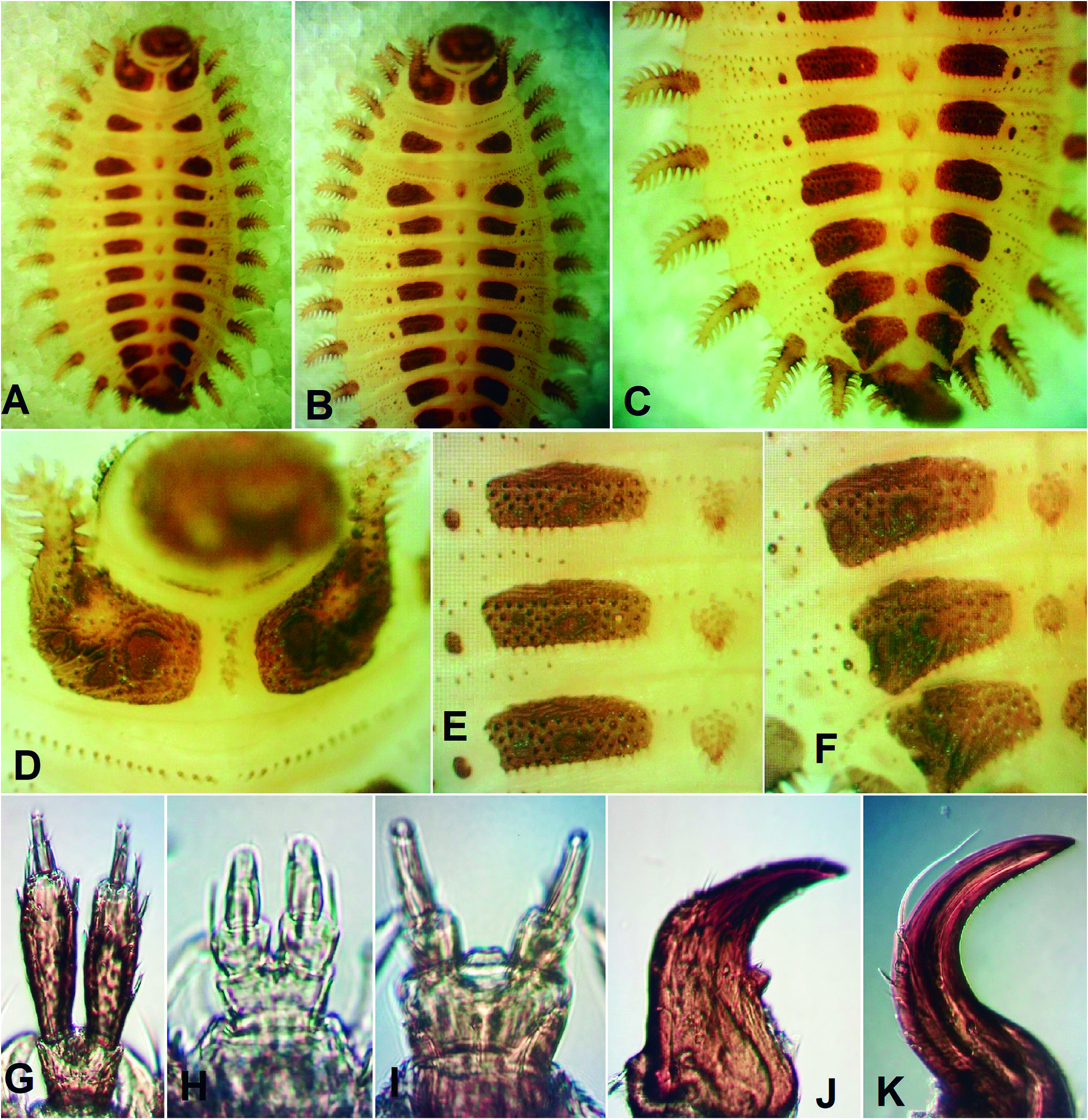
( Figs. 1 View Figure 1 , 2 View Figure 2 , 3 View Figure 3 A-F, 3I, 3K, 5C-F, 6B)
Type material: HOLOTYPE: ♀: AUSTRALIA: “QLD: 23.707°S × 141.147°E 6.3 km N of Diamantina NP HQ , 120 m, 03Oct2011, Monteith & Turco, ex bark of gidgee trees, 39461 /adult ♀ emerged 17-18.xi.2011 ” ( QMB, Reg. No. T245515 ). The dried last larval and pupal exuviae of the holotype are card-mounted and stored with the holotype. GoogleMaps
Differential diagnosis: Distinguishing this species from the seven other described forms currently included in the genus on the basis of a single female is difficult, since most key characters are based on males, and the two sexes in this genus appear to differ considerably, at least in size, shape and antennal structure.The B. cleidecostae female at least differs from females of B. fulva with respect to features listed in Table 1. It also differs from females of B. lampyroides and an undescribed species from Cyprus (Petrzelkova et al., 2017) with respect to the size and pronotal shape (Characters 1 and 2), which are similar to those features in B. fulva .
Description (Adult female): Length 3.8 mm from anteri- or margin of head to apex of elytra (excluding extended abdominal apex); combined pronotal and elytral length 1.90 times greatest elytral width. Color yellowish-brown; dorsal vestiture of fine, suberect setae. Head transverse, more or less declined and deeply inserted into prothorax. Eye ( Figs. 1 View Figure 1 E-F) 0.32 times as long as head width behind eyes, finely facetted. Antennal insertions completely exposed, located in large, saucer-like impressions separated by slightly more than one diameter. Frontal area declined; clypeus very short and broad, anteriorly emarginate; labrum about 0.5 times as long as wide, 0.35 times as wide as clypeus and broadly rounded at apex. Antennae ( Fig. 1E View Figure 1 ) about 1.4 times as long as head width behind eyes; scape 1.25 times as long as wide, pedicel about as long as wide, both subglobular but slightly wid- er apically; antennomere 3 about 1.2 times as long as wide and 1.2 times as long as 4; 5-10 subequal in length, gradually shorter and broader and expanded apically; antennomere 11 twice as long as 10, 1.25 times as long as wide and apically subacute. Mandibles broad at base, unidentate. Apical maxillary and labial palpomeres elongate, slender, widest at middle and narrowly rounded or truncate at apex. Pronotum ( Figs. 1 View Figure 1 A-C) 0.49 times as long as its greatest width (across posterior angles) and 0.54 times as long as its width at about middle; sides gradually converging anteriorly; lateral pronotal carinae in dorsal view visible posteriorly but concealed anteriorly by sides of disc; posterior pronotal angles strongly produced posterolaterally and acute; sublateral carina extending from apex of posterior angle almost to middle of disc and accompanied mesally by a short, weak, longitudinal groove ( Fig. 1F View Figure 1 ); anterior edge of pronotum subtruncate; posterior edge weakly triemarginate, with two broad lateral emarginations flanking a relatively narrow prescutellar emargination. Disc coarsely and densely punctate with punctures separated by less than half a diameter; setae moderately long and fine. Prosternum ( Fig.1C View Figure 1 ) about 1.9 times as long as mid length of procoxal cavity, with short, broad chin piece at anterior end; prosternal process about 0.50 times as wide as mid length of coxal cavity, parallel sided, extending to posterior edges of coxae, with narrowly rounded apex. Procoxal cavities widely open; postcoxal (hypomeral) processes very short and angulate; protrochantin concealed by coxal cowling. Scutellar shield 0.92 times as long as wide, weakly curved anteriorly, broadly rounded posteriorly. Elytra ( Figs. 1 View Figure 1 A-B) 1.48 times as long as greatest width (at about middle); humeri well-developed but somewhat flattened; lateral margins well-developed but concealed anteriorly by humeri; disc with a series of shallow, longitudinal grooves; punctation similar to that on pronotum, but less regular and bearing moderately long, fine setae; apices independently rounded. Elytral epipleura narrow and more or less complete. Hindwings (as seen through elytral cuticle) well-developed, with relatively small apical field, elongate radial cell with oblique base and obtuse inner angle, short, slightly oblique cross-vein r3 and well-developed apical spur (continuation of MP₁ ₊ ₂); the medial field, however, is folded under, concealing the condition of MP₃ ₊ ₄, Cu, AA and AP. Mesocoxal cavities ( Fig. 1D View Figure 1 ) sep- arated by a distance subequal to the greatest longitudinal diameter of a cavity, partly closed laterally by mesanepisternum and mesepimeron; mesotrochantin concealed; mesoventrite with mesal cavity extending almost to posterior edge, which is very shallowly emarginate. Metaventrite, excluding anterior lobe, about 0.50 times as long as wide,moderately convex; discrimen 0.40 times as long as ventrite; posterior edge between metacoxae deeply, broadly emarginate. Metanepisternum 3.33 times as long as wide; more or less parallel-sided, but with curved carina extending from near anteromesal edge to lateral edge at about middle. Metacoxae ( Fig. 1D View Figure 1 ) separated by a distance equal to longest longitudinal diameter of coxa; coxal plates weakly developed but complete to lateral edges. All trochanters at least slightly elongate, trochanterofemoral joints varying from very slightly oblique (fore legs) to strongly oblique (hind legs); legs ( Fig. 1D View Figure 1 ) relatively short and slender, with femora, tibiae and tarsi all about equal in length; tarsi and pretarsal claws simple. Abdominal ventrite 1 ( Figs. 1 View Figure 1 C-D) about 0.75 times as long as 2, 3 or 4; ventrite 5 about 1.5 times as long as 4 and very broadly rounded; posterior portions of ventrites 1-4 membranous. Most abdominal tergites lightly sclerotized, but those of segments VII and VIII more heavily sclerotized ( Figs. 1 View Figure 1 A-B). Ovipositor ( Fig. 1A View Figure 1 ) short, broad and lightly sclerotized (possibly not yet fully pigmented), with gonocoxites broadly rounded at apex and gonostyli slender and parallel-sided.
Larval specimens examined: AUSTRALIA. Queensland: 8 km N of Diamantina Lakes Homestead ( 23°42’25”S, 141°08’48”E), 21.xi.2002, under logs and in litter of stunt- ed acacia on marly limestone, R. Crookshanks,2examples ( Fig. 2 View Figure 2 ) ( QMB) GoogleMaps ; 6 km N of Diamantina Lakes National Park HQ ( 23.7070S, 141.1467E), 120 m, 03.x.2011, under bark flakes of Gidgee trees ( Acacia cambagei R. T. Baker ), G.B. Monteith & F. Turco, 2 examples and one partial exuvia ( Figs. 3I, 3K View Figure 3 , 5C View Figure 5 ) ( QMB) GoogleMaps ; South Australia: Flinders Range, Arkaroola Sanctuary, Stubbs Waterhole , 22.x.2007, under fibrous bark of Eucalyptus, M. Anstis & W. Grimm, 1 example ( Figs. 3 View Figure 3 A-F, 6A) ( AMS: K379869 ) ; Western Australia: Winjana [sic, = Windjana)] Gorge , 100 m, 17.x.1962, E.S. Ross & D. Cavagnaro, 2 examples ( CAS) .
Larval differential diagnosis: The larva of B. cleidecostae was shown by Costa et al. (2006) and Petrzelkova et al. (2017) to differ from those of B. fulva , B. moronei , B. lampyroides and an unnamed larva from Cyprus by the pedunculate stemmata, absence of frontal arms (ecdy-
sial lines) and gular sutures, mandible with a retinaculum ( Fig. 3J View Figure 3 ), and undivided anterior lobe of the second abdominal segment (Costa et al., 2006, figs. 74-86). In the present study a number of additional features were found to distinguish this new species from B.fulva , based on comparisons of dissected specimens and illustrations in Costa et al. (2006), and from B. lampyroides based on illustrations in Costa et al. (2006) and Klausnitzer (2009); these are summarized in Table 2. Among the fossil specimens, that of B. moronei Branham has unusual maxillary and labial palps resembling those in B. fulva but with the first labial palpomere expanded towards apex, while that of an unnamed species from Cyprus (Petrzelkova et al., 2017) closely resembles the larva of B. lampyroides from southern India, as illustrated in Costa et al. (2006); whether that larva is conspecific with those illustrated by Klausnitzer (2009) from Goa is uncertain. It is interesting to note that the greatly expanded maxillary and labial palps of late instar B. fulva and B. moronei are not present in first instar larvae of the former species, based on specimens collected in southern California.
Larval description ( Figs. 2-3 View Figure 2 View Figure 3 ): Length of largest larva (Arkaroola, SA) about 8.8 mm including head and tail spine; major portion of body 5.8 mm long and about 1.2 times as long as wide, including lateral branched lobes, which occur on all body segments excluding IX and X. Body flattened; upper surfaces completely covered with numerous protuberances (small tubercles to long, narrow tubules) bearing a variety of modified scale-like setae and sclerotized plates of varying size and extent. Head dorsally about 0.9 times as long as wide and ventrally 1.45 times as long as wide, sides more or less parallel behind stemmata, then slightly narrowed anteriorly. Ecdysial lines consisting of very short epicranial stem but no frontal arms; an additional pair of ecdysial lines or lines of weakness extending laterally from the epicranial stem, separating a pair of postepicranial plates. Dorsal and ventral surface densely clothed with narrow, tubular projections bearing complex setae ( Fig. 2B View Figure 2 ). Stemmata relatively large and pedunculate. Frontoclypeal area just in front of stemmata slightly declined; frontoclypeal suture absent.Anterior edge of clypeus weakly emarginate. Antenna ( Fig. 2B View Figure 2 ) about half as long as head width, extending anteriorly but slightly tilted laterally; antennomere 1 short, transverse and simple, 2 about 8.2 times as long as 1 and 2.6 times as long as wide, narrow at base, gradually expanded to apical third, then narrowed to broadly rounded apex, its dorsal and lateral surfaces covered with complex setae, but without tubular projections and its ventral surface smooth; antennomere 3 and sensorium minute, lying side by side at the apex of antennomere 2, with the sensorium subconical and about 0.75 times as long as antennomere 3. Labrum subquadrate, its apex concave with pair of paramedian setiferous tubules located posterior to anterolateral ones. Mandible ( Fig. 3J View Figure 3 ) 1.85 times as long as wide, widest at base; outer edge straight to middle then abruptly curved mesally; apex unidentate; incisor edge with small, acute retinaculum; and base slightly expanded but without any obvi- ous armature; internal channel present. Ventral mouthparts deeply retracted; hypostomal cavity with straight, posteriorly converging sides and subtruncate posterior edge. Cardines completely fused together forming a trapezoidal plate; stipites elongate and slender, converging posteriorly but slightly separated by postmentum; lacinia lightly sclerotized, curved and narrowly acute at apex; galea small, articulated with apical seta; maxillary palp 4-segmented, ratio of palpomere lengths 1.07: 1.00: 1.36: 1.14; first palpomere 0.67 times as long as wide, with setose tubule attached to inner edge; palpomeres 2-4 each narrower than the one preceding it; second palpomere 0.8 times as long as wide; third 1.5 times as long as wide; terminal palpomere 3.8 times as long as wide. Prementum ( Fig. 3G View Figure 3 ) slightly transverse; labial palps separated by a distance about as wide as a basal palpomere; apical palpomere about 1.5 times as long as basal one and distinctly narrower. Gular sutures absent. Dorsal surfaces ( Figs. 3 View Figure 3 A-F) of all thoracic and abdominal segments with scattered, minute setiferous tubercles and anterior edges of meso- and metaterga each with transverse row of similar tubercles; cervical region with pair of slender, oblique sclerites; thoracic terga and abdominal terga 1-8 each with a pair of darkly pigmented plates extending from near the midline usually to the lateral fourth of the tergum; plates of protergum ( Fig. 3D View Figure 3 ) irregularly quadrate, each anterolaterally contiguous with an anteriorly projecting branched lobe; plates of the meso- and metaterga ( Figs. 3 View Figure 3 A-B) more or less teardrop-shaped, subacute mesally and broadly rounded laterally, more widely separated at middle than on protergum; plates of abdominal terga 1-6 ( Figs. 3 View Figure 3 E-F) narrowly quadrate, broadly rounded or truncate at each end and more widely separated at midline; plates of tergum 7 shorter, broader and more narrowly separated at midline, those on tergum 8 even shorter, broader and contiguous at midline. Mesal interspaces between paired plates lightly pigmented, that on protergum with a small cluster of setiferous tubules anteriorly and a larger one posteriorly, that on mesotergum with a small circular prominence covered with setiferous tubules and a small pair of papillae, that on metatergum with the largest somewhat conical prominence and a darker lobe posteriorly bearing paired papillae, and those on abdominal terga 1-7 with gradually smaller prominences with similar lobes bearing paired papillae. Lateral apigmented areas of abdominal terga 1-4 each with a small, circular dark plate mesally and a small spiracle on a short tube laterally; on tergum 5 same area with circular plate partly fused to median plate and on terga 6 and 7 plate entirely fused to median plate; spiracles on 5-7 similar to those anteriorly, but that on tergum 8 fused to median plate. All thoracic terga and abdominal terga I-VIII with two pairs of lateral lobes ( Figs. 2A View Figure 2 , 3 View Figure 3 A-F, 6B); both pairs on thoracic segments and posterior pair on abdominal segments similarly shaped, large, elongate, moderately sclerotized and horizontally branched,with more than 10 branches, each bearing a modified seta; anterior lobes on all abdominal segments highly reduced, lightly sclerotized, bearing one or a few setiferous tubules, and often difficult to see. Anterior branched lobe on protergum projecting anteriorly, narrowed and curved laterally at apex, shorter than posterior lobe, which is straight and projecting anterolaterally; branched lobes on meso- and metaterga similar in length, narrowed apically and curved posteriorly; posterior lobes on abdominal terga I-VII similar in shape and curved posteriorly, but those on VIII slightly shorter, straight and projecting posterolaterally. Tergum IX forming a flexible tail spine about 3.4 times as long as wide, slightly curved dorsally, widest at base and narrowing to subacute apex, heavily sclerotized, and armed with a series of setiferous tubules, the first 4 lateral longest and the remainder shorter and varying in size, with lateral and ventral tubules longer than those on dorsum,except for a medial longitudinal row of longer dorsal tubules. Ventral surfaces, excluding larger lateral branched lobes and tail spine, lightly sclerotized or membranous, except for cervicosternum,which has paired subtriangular plates narrowly separated at midline, sterna VIII and IX, each of which bears a pair of small, circular plates, mesothoracic spiracular tubes and legs. Prosternum lightly pigmented, but without distinct plates; meso- and metathorax mainly membranous, with precoxal sclerites lightly pigment- ed. Spiracular tubes located on anterolateral portions of mesosternum, close to edge of prosternum, each one more or less conical, well sclerotized and facing laterally. Legs relatively heavily sclerotised and armed with short spicules; fore leg slightly shorter than midleg, which is 0.67 times as long as width of mesothorax excluding lateral processes, with coxa short and subconical, trochanter and femur separated by a diagonal suture and the two together 1.14 times as long as tibiotarsus; pretarsus 0.43 times as long as tibiotarsus with paired setae lying side by side near base. Most abdominal sterna membranous or very lightly sclerotized but clothed with a series of scattered, very fine tubercles.
Etymology: Named for Professor Cleide Costa in honour of her numerous contributions to Entomology and particularly her work on beetle larvae.
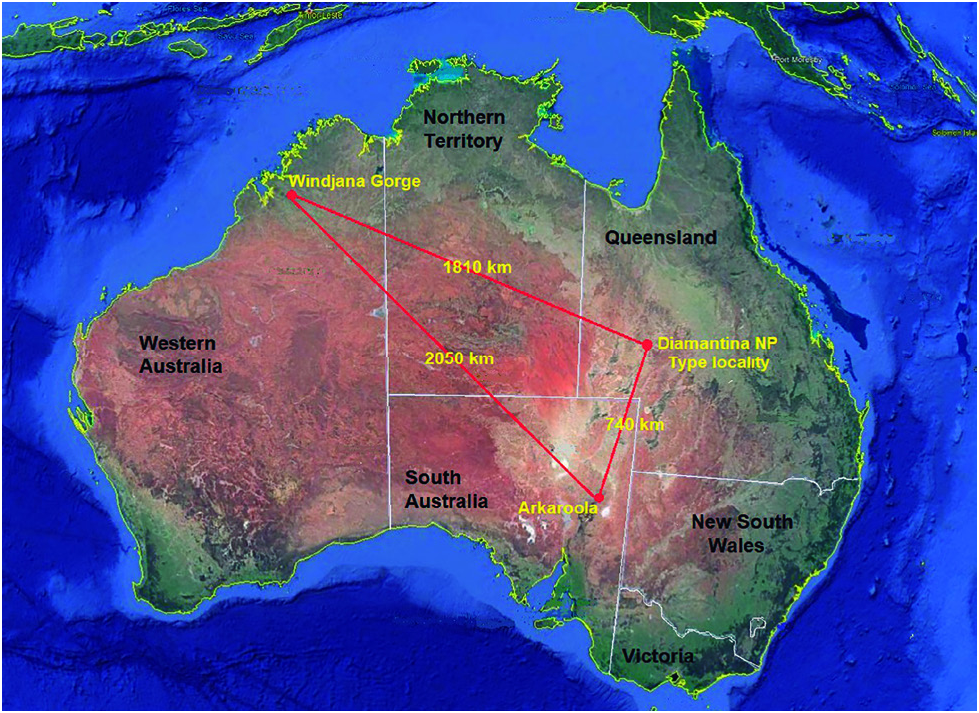
Distribution and habitat: Larval specimens of Brachypsectra cleidecostae sp. nov. reported on in this paper have been collected on four occasions from three very widely separated localities ( Fig. 4 View Figure 4 ) in three different Australian states ( Western Australia, Queensland and South Australia) over a time span of 49 years. The wide spread of these localities, coupled with the apparent rarity and/or crypticity of specimens and the fact that all collections appear to be conspecific, indicates that B. cleidecostae is probably widespread through the arid parts of the continent. The fact that the habitat and climate is quite different at each of the three sites also suggests that the species does not have habitat restrictions on its distribution. Windjana Gorge lies at 125 m elevation, is dominated by low eucalypt woodland and has 600-800 mm annual rainfall, almost all rain falling during the summer monsoon period, leaving most of the year semi-arid. Diamantina Lakes NP ( Figs. 5 View Figure 5 A-B) is at 120 m elevation with sparse desert shrubland and scattered, stunted trees, and only 200-300 mm of annual rainfall, which also is largely dependent on the summer monsoon season. Stubbs Waterhole at Arkaroola ( Fig. 6A View Figure 6 ) is at 220 m elevation, with annual rainfall of 250 mm, mostly falling in the winter months, in a rocky, mountainous landscape with scattered eucalypt woodland.
Circumstances of collection in Australia: The first collection was by CAS entomologists, Ed Ross and David Cavagnaro, who took two larvae at Windjana Gorge in the arid NW of Western Australia in October 1962. No collection method is given but since leader Ed Ross’s main interest was in Embioptera, which occur frequently under bark, they may have been taken in that situation. These specimens were the basis of the first notice and illustration of the family in Australia by Lawrence & Britton (1991).
The second Australian collection was by Roy Crookshanks, an amateur mollusc enthusiast who regularly sent land snails to the QMB malacology collection. Among a batch of small snails sent in 2002 from the remote desert reserve of Diamantina Lakes National Park were two dried larvae ( Fig. 2 View Figure 2 ) of B. cleidecostae , included because of their unusual form. Crookshanks reported (pers. comm.) that they were found while searching un- der logs and in leaf litter in a patch of stunted acacia trees, and he gave the exact GPS coordinates of the site. Nine years later, in October 2011, QMB entomologists Federica Turco and co-author Geoff Monteith returned to the exact spot and found a small dense patch of gidgee ( Acacia cambagei R.T. Baker) and bauhinia ( Lysiphyllum sp.) trees about 25 m in diameter in an otherwise harsh desert shrubland landscape ( Fig. 5A View Figure 5 ). Gidgee trunks have many layers of hard, persistent bark flakes ( Fig. 5B View Figure 5 ) with numerous narrow interspaces which seemed suitable for brachypsectrid larvae. In several hours of systematic levering off of bark flakes with screwdrivers, one large ( 6 mm) and one small ( 3 mm) larvae were found on October 3, both by Turco ( Fig. 5C View Figure 5 ). The smaller larva was free living but the larger was enclosed in an oval pupal shelter formed of stiff, thick, white, silken mesh joining two bark surfaces ( Figs. 5 View Figure 5 E-F), identical to the shelters described for the north American Brachypsectra fulva by Fleenor &Taber (1999) and illustrated by a photograph of a laboratory reared specimen taken by Margaret Thayer in Costa et al. (2006). The two specimens were taken live to Brisbane where, after photography during which the small larva was lost, the large larva built a new shelter between bark and tissue paper in a petri dish on about 8-9 October. The silken mesh was incomplete and of much finer threads than the original. The larva remained in a rigid S-shaped posture, identical to that shown in the Thayer photograph of B. fulva , until 28 October when it pupated. The pupa remained partly sheathed in the stiff larval exuvia and moved actively when touched. On the night of November 17 a female emerged ( Fig. 5D View Figure 5 ). It had a greatly distended abdomen and exuded a quantity of thick creamy material. It remained inactive and died after 2-3 days. This specimen is the holotype of B. cleidecostae sp. nov. Its pupal period of 3 and a half weeks was considerably less than the six weeks recorded for B. fulva in North America ( Fleenor & Taber, 1999). The collection of these Diamantina NP specimens was informally recorded by Monteith & Turco (2011).
The fourth collection from Australia was at Stubbs Waterhole, 6 km east of the homestead of Arkaroola private conservation sanctuary ( Fig. 6A View Figure 6 ),at the northern end of the Flinders Ranges in South Australia. Herpetologists Marion Anstis and Wendy Grimm, associates of AMS, took a large ( 8.8 mm) larva ( Fig.6B View Figure 6 ) in October 2007 while peeling fibrous bark from a eucalyptus tree ( Fig. 6A View Figure 6 ) in search of insect food for captured frogs and geckos. The specimen was photographed while alive, then preserved and returned to AMS where it was recognised as Brachypsectridae . Photographs were published informally by Grimm & Grimm (2016).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |