
Synergus japonicus Walker, 1874
|
publication ID |
https://doi.org/ 10.11865/j.zs.20140406 |
|
publication LSID |
lsid:zoobank.org:pub:053DEE6B-462D-483A-ABCB-7FF46C6BC8B1 |
|
DOI |
https://doi.org/10.5281/zenodo.4617476 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA5B61-EF64-742D-FF4E-FE06FB98FD40 |
|
treatment provided by |
Carolina |
|
scientific name |
Synergus japonicus Walker, 1874 |
| status |
|
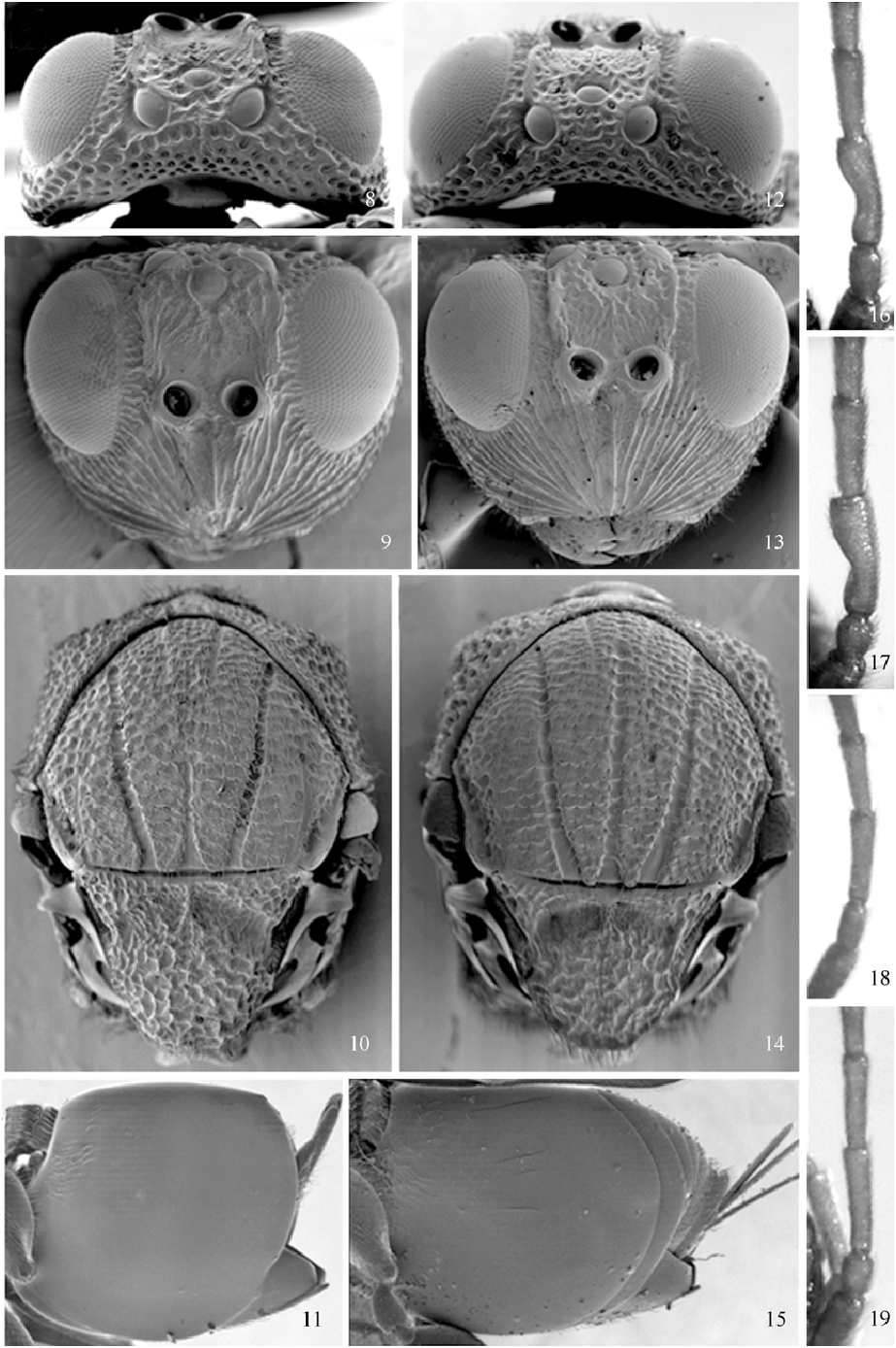
Synergus japonicus Walker, 1874 New record to China ( Figs 12–15, 17, 19 View Figs 8–19 )
Type material examined. Holotype ♀, deposited in NHM, with following labels: “2b2” (white label handwritten), “ Japan, Coll. F. Walker. 1913-71” (white label impressed), “Type” (round and white impressed label with red colour in the margin), “ Synergus japonicus ” (white label handwritten), “ B. M. Type Hym., 7.109” (white label).
Additional material. 5♂, 5♀, Chohabaru, Oita, Japan, ex. Galls of the unisexual generation of Andricus kashiwaphilus , April 1986, leg. Y. Abe. 2♀, Yamanashi, Japan), June 1950, ( Synergus watensis (Shingi) (= jezoensis U + S) det unknown-48), letter “B”, USNM 2017203. 1♀, Yamanashi, Japan), June 1950, letter “B”, USNM 2017203. 3♀, Boshi, Youxian, Hunan, China, 26 April 1993) April 1993, ex. Andricus mukaigawae on Quercus serrata , leg. Zhi-Wei Liu.
Diagnosis. The species is very similar to S. hayneanus in morphology and color, but differs from the latter by a combination of the following characters: (1) gena slightly expanded laterally; (2) transverse carinae of mesoscutum heavier and more spaced apart; (3) presence of central carinae between lateral carinae of propodeum (absent in S. hayneanus ); (4) extent of pilosity of fused segments 2+3, with a patch of hair in S. japonicas (with a row of hairs in S. hayneanus ); (5) the extended band of micropunctures of metasomal syntergite incomplete, slightly short of reaching the ventral margin; (6) the shape of F1 and the chromatic characteristics of head in males different (details in description below).
Redescription. Length. Female 2.6–3.8 mm (one specimen 2.3), male 2.2–2.9 mm.
Color. Female. Head black except a small area dark red around the mouth and near the compound eyes; mandibles yellow, the first tooth red and the rest black; antennae testaceous: mesosoma black; legs dark, coxae black or dark brown, femur brown to dark brown, tibia and tarsus lighter; wings hyaline, veins yellowish; metasoma brown or reddish brown. Male. Head basically amberous, dorsally black; coxae I, II and sometimes the coxae III yellowish-brown; veins darker.
Head ( Figs 12–13 View Figs 8–19 ). In frontal view, about 1.3 times as wide as height, and in dorsal view about twice as wide as long. Face with strong carinae radiating from clypeus, median carina not raised. Malar space slightly more than 0.5 times the height of compound eye. Transfacial line only slightly shorter than height of compound eye. Toruli diameter equal to the distance between the margin of torulus and margin the compound eye (sligthly longer in males), and distance between toruli sligtly smaller than its diameter (narrower in males). Frons densely punctate. Frontal carina present. Vertex and occiput strongly punctate with interspaces coriaceous. POL: LOL: OOL = 8: 4: 3 (in males 9: 4: 4), diameter of occelus 4 in proportion (3 in males). Gena slightly expanded behind eye, punctate with interspaces coriaceous.
Antenna. Female ( Fig. 19 View Figs 8–19 ). Pedicellum slightly longer than wide; F1 cylindrical, slightly broader distally; antennal formula 9: 4: 11: 10: 10: 10: 8: 8: 7: 6: 5: 5: 5: 9. Male ( Fig. 17 View Figs 8–19 ). F1 curved and more expanded distally than basally; antennal formula 6: 3: 11: 9: 8: 7: 7: 6: 6: 5: 5: 5: 4: 4: 4.
Mesosoma ( Fig. 14 View Figs 8–19 ). Subsquare in lateral view. Pronotum robust with lateral carina, laterally rugose, and strongly carinate inferiorly. Mesoscutum strongly transversely carinate with interespaces coriaceous (much less obvious and hardly impressed in males). Notauli wide, strongly impressed, percurrent; medial scutum line long, almost percurrent. Mesopleuron entirely longitudinally carinated. Scutellum rugose, interspaces coriaceous; scutellar foveae large and oblique, separated by a incomplete median carina, and coriaceous and pubescent; lateral propodeal carinae straight, delimiting a squared pubescent and coriaceous area, with a median longitudinal carina and a few transverse carinae.
Forewings. Pubescent. Radial cell around 2.2 times as long as wide. Areolet inconspicous (more distinct in males); vein Rs curved; vein Rs+M indistinct.
Metasoma ( Fig. 15 View Figs 8–19 ). Longer than mesosoma. Fused segment 2+3 strongly incised dorso-distally with a patch of setae antero-laterally; apical patch of micropunctures extending to 1/3 length of metasoma, slightly short of reaching the ventral margin of syntergite; subsequent tergites punctured; hypopygium shinning, without setae and punctures only with a few puncures apically. Ventral spine of hypopygium very short.
Variation. One small female specimen deposited in USNM (collected June 1950) has areolet missing, coxae similarly colored as legs, and the reddish area surrounding mouth is more extended in face and genae.
Distribution. China (Hunan), Japan.
Biology. The species, which was referred to as Synergus japonicus type A by Abe (1990), is arrhenotokous and bivoltine (Abe, 1990), and may have a third generation in the summer (Abe, 1992). The first generation emerges in May from the galls of the unisexual generation of Andricus mukaigawae and the second generation emerges in June from the galls of the bisexual generation of Biorhiza nawai . Two consecutive emergences from B. nawai may be observed sometimes in the spring or a prolonged emergence may occur from spring throughout summer. Galls of A. mukaigawai infected with S. japonicus inquilines were smaller compared with those, infected with S. gifuensis and the inquiline larval cells are separated by a membranous wall inside the host gall (Abe, 1990, 1992). Although there were evidences suggesting that S. japonicas females may oviposit in host galls already infected with S. gifuensis , the two species are reproductively isolated from each other by ethological barrier (Abe 1992). Collection data (see above) indicate that the species can also exploit the galls of the unisexual generation of Andricus kashiwaphilus .
| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |